Patoloogias on üheks kõige sagedamini häiritud ainevahetuse tüübiks vesi-sool. See on seotud vee pideva liikumisega ja mineraalid organismi väliskeskkonnast sisemisse ja vastupidi.
Täiskasvanu kehas moodustab vesi 2/3 (58-67%) kehakaalust. Umbes pool selle mahust on koondunud lihastesse. Veevajadus (inimene saab päevas kuni 2,5–3 liitrit vedelikku) kaetakse selle tarbimisega joogina (700–1700 ml), toiduga ette nähtud vee (800–1000 ml) ja vesi , moodustub organismis ainevahetuse käigus - 200--300 ml (100 g rasvade, valkude ja süsivesikute põletamisel tekib vastavalt 107,41 ja 55 g vett). Endogeenne vesi suhteliselt suurel hulgal sünteesitakse rasvade oksüdatsiooniprotsessi aktiveerimisel, mida täheldatakse mitmesugustes, peamiselt pikaajalistes stressitingimustes, sümpaatilise-neerupealise süsteemi ergutamisel, mahalaadimise dieediteraapias (kasutatakse sageli rasvunud patsientide raviks).
Pidevalt esinevate kohustuslike veekadude tõttu jääb vedeliku sisemaht organismis muutumatuks. Need kaotused hõlmavad neerude (1,5 l) ja ekstrarenaalset kahju, mis on seotud vedeliku vabanemisega seedetrakti kaudu (50–300 ml), Hingamisteed ja nahk (850-1200 ml). Üldjuhul on kohustuslike veekadude maht 2,5-3 liitrit, mis sõltub suuresti organismist eemaldatud mürkainete hulgast.
Vee roll eluprotsessides on väga mitmekesine. Vesi on paljude ühendite lahusti, mitmete füüsikalis-keemiliste ja biokeemiliste transformatsioonide otsene komponent, endo- ja eksogeensete ainete transportija. Lisaks täidab see mehaanilist funktsiooni, nõrgendades sidemete, lihaste, liigeste kõhrepindade hõõrdumist (hõlbustades seeläbi nende liikuvust) ja osaleb termoregulatsioonis. Vesi säilitab homöostaasi, mis sõltub plasma osmootse rõhu suurusest (isoosmia) ja vedeliku mahust (isovoleemia), happe-aluse oleku reguleerimise mehhanismide toimimisest, temperatuuri püsivust tagavate protsesside toimumisest. (isotermia).
Inimkehas esineb vesi kolmes peamises füüsikalises ja keemilises olekus, mille järgi nad eristavad: 1) vaba ehk liikuvat vett (moodustab põhiosa rakusisesest vedelikust, samuti verest, lümfist, interstitsiaalsest vedelikust); 2) vesi, mis on seotud hüdrofiilsete kolloididega ja 3) konstitutsiooniline, sisaldub valkude, rasvade ja süsivesikute molekulide struktuuris.
70 kg kaaluva täiskasvanud inimese kehas on vaba vee ja hüdrofiilsete kolloididega seotud vee maht ligikaudu 60% kehamassist, s.o. 42 l. Seda vedelikku esindavad rakusisene vesi (see moodustab 28 liitrit ehk 40% kehamassist), mis moodustab rakusisese sektori, ja rakuväline vesi (14 liitrit ehk 20% kehamassist), mis moodustab rakuvälise sektori. Viimase koostis sisaldab intravaskulaarset (intravaskulaarset) vedelikku. Selle intravaskulaarse sektori moodustavad plasma (2,8 l), mis moodustab 4-5% kehamassist, ja lümf.
Interstitsiaalne vesi sisaldab korralikku rakkudevahelist vett (vaba rakkudevaheline vedelik) ja organiseeritud rakuvälist vedelikku (moodustades 15–16% kehamassist ehk 10,5 liitrit), s.o. sidemete, kõõluste, fastsia, kõhre jne vesi. Lisaks sisaldab ekstratsellulaarne sektor vett, mis asub mõnes õõnsuses (kõhu- ja pleura õõnsus, perikardis, liigestes, ajuvatsakestes, silmakambrites jne), samuti seedetraktis. Nende õõnsuste vedelik ei osale aktiivselt ainevahetusprotsessides.
Vesi Inimkeha ei seisa oma erinevates osakondades, vaid liigub pidevalt, vahetades pidevalt teiste vedeliku sektorite ja väliskeskkonnaga. Vee liikumine on suuresti tingitud seedemahlade eraldumisest. Seega koos süljega koos kõhunäärme mahlaga saadetakse soolestikku umbes 8 liitrit vett päevas, kuid see vesi imendub madalamates piirkondades. seedetrakt peaaegu kunagi ei kao.
Elutähtsad elemendid jagunevad makrotoitaineteks ( igapäevane vajadus>100 mg) ja mikroelemendid (päevane vajadus<100 мг). К макроэлементам относятся натрий (Na), калий (К), кальций (Ca), магний (Мg), хлор (Cl), фосфор (Р), сера (S) и иод (I). К жизненно важным микроэлементам, необходимым лишь в следовых количествах, относятся железо (Fe), цинк (Zn), марганец (Мn), медь (Cu), кобальт (Со), хром (Сr), селен (Se) и молибден (Мо). Фтор (F) не принадлежит к этой группе, однако он необходим для поддержания в здоровом состоянии костной и зубной ткани. Вопрос относительно принадлежности к жизненно важным микроэлементам ванадия, никеля, олова, бора и кремния остается открытым. Такие элементы принято называть условно эссенциальными.
Kuna organismis saab ladestuda palju elemente, kompenseeritakse kõrvalekalle päevasest normist õigeaegselt. Kaltsium apatiidi kujul ladestub luukoes, jood ladestub türeoglobuliini osana kilpnäärmes, raud ladestub ferritiini ja hemosideriini koostises luuüdis, põrnas ja maksas. Maks toimib paljude mikroelementide hoiukohana.
Mineraalide ainevahetust kontrollivad hormoonid. See puudutab näiteks H2O, Ca2+, PO43- tarbimist, Fe2+, I- sidumist, H2O, Na+, Ca2+, PO43- eritumist.
Toidust omastatavate mineraalainete hulk sõltub reeglina organismi ainevahetusvajadustest ja mõnel juhul ka toiduainete koostisest. Kaltsiumi võib pidada toidu koostise mõju näiteks. Ca2+ ioonide imendumist soodustavad piim- ja sidrunhape, fosfaadi ioon, oksalaadi ioon ja fütiinhape aga pärsivad kompleksi moodustumise ja halvasti lahustuvate soolade (fütiin) moodustumise tõttu kaltsiumi imendumist.
Mineraalipuudus pole haruldane nähtus: see tekib erinevatel põhjustel, näiteks monotoonse toitumise, seedimishäirete ja erinevate haiguste tõttu. Kaltsiumipuudus võib tekkida raseduse ajal, samuti rahhiidi või osteoporoosi korral. Klooripuudus tekib Cl-ionide suure kaotuse tõttu koos tugeva oksendamisega.
Toiduainete ebapiisava joodisisalduse tõttu on mitmel pool Kesk-Euroopas levinud joodipuudus ja struuma. Magneesiumipuudus võib tekkida kõhulahtisuse või alkoholismi korral monotoonse toitumise tõttu. Mikroelementide puudumine kehas väljendub sageli vereloome rikkumises, st aneemias.
Viimases veerus on loetletud funktsioonid, mida need mineraalid kehas täidavad. Tabeli andmetest on näha, et peaaegu kõik makrotoitained toimivad organismis struktuurikomponentidena ja elektrolüütidena. Signaalifunktsioone täidavad jood (jodotüroniini osana) ja kaltsium. Enamik mikroelemente on valkude kofaktorid, peamiselt ensüümid. Kvantitatiivselt on organismis ülekaalus rauda sisaldavad valgud hemoglobiin, müoglobiin ja tsütokroom, aga ka üle 300 tsinki sisaldava valgu.
Vee-soola ainevahetuse reguleerimine. Vasopressiini, aldosterooni ja reniin-angiotensiini süsteemi roll
Vee-soola homöostaasi peamised parameetrid on osmootne rõhk, pH ning rakusisese ja rakuvälise vedeliku maht. Nende seadete muutmine võib muutuda vererõhk, atsidoos või alkaloos, dehüdratsioon ja tursed. Peamised vee-soola tasakaalu reguleerimises osalevad hormoonid on ADH, aldosteroon ja kodade natriureetiline faktor (PNF).
ADH ehk vasopressiin on 9 aminohappest koosnev peptiid, mis on ühendatud ühe disulfiidsillaga. See sünteesitakse hüpotalamuses prohormoonina, seejärel kantakse hüpofüüsi tagumise osa närvilõpmetesse, kust eritub sobiva stimulatsiooniga vereringesse. Liikumine piki aksonit on seotud spetsiifilise kandjavalguga (neurofüsiin)
ADH sekretsiooni põhjustav stiimul on naatriumioonide kontsentratsiooni tõus ja rakuvälise vedeliku osmootse rõhu tõus.
ADH jaoks on kõige olulisemad sihtrakud distaalsete tuubulite ja neerude kogumiskanalite rakud. Nende kanalite rakud on suhteliselt vett mitteläbilaskvad ning ADH puudumisel uriin ei kontsentreerita ning seda võib väljutada üle 20 liitri ööpäevas (norm 1-1,5 liitrit ööpäevas).
ADH jaoks on kahte tüüpi retseptoreid, V1 ja V2. V2 retseptorit leidub ainult neeruepiteelirakkude pinnal. ADH seondumine V2-ga on seotud adenülaattsüklaasi süsteemiga ja stimuleerib proteiinkinaasi A (PKA) aktivatsiooni. PKA fosforüleerib valke, mis stimuleerivad membraanivalgu geeni akvaporiin-2 ekspressiooni. Aquaporin 2 liigub apikaalsele membraanile, siseneb sellesse ja moodustab veekanalid. Need tagavad rakumembraani selektiivse vee läbilaskvuse. Veemolekulid difundeeruvad vabalt neerutuubulite rakkudesse ja sisenevad seejärel interstitsiaalsesse ruumi. Selle tulemusena imendub vesi neerutuubulitest tagasi. V1 tüüpi retseptorid paiknevad silelihaste membraanides. ADH interaktsioon V1 retseptoriga viib fosfolipaasi C aktiveerimiseni, mis hüdrolüüsib fosfatidüülinositool-4,5-bifosfaati IP-3 moodustumisega. IF-3 põhjustab Ca2+ vabanemise endoplasmaatilisest retikulumist. Hormooni toime tulemusena V1 retseptorite kaudu toimub veresoonte silelihaskihi kokkutõmbumine.
Hüpofüüsi tagumise osa talitlushäiretest põhjustatud ADH defitsiit, samuti hormonaalse signaalisüsteemi häired võivad põhjustada diabeedi insipidus'e arengut. Diabeedi insipiduse peamine ilming on polüuuria, s.o. suures koguses madala tihedusega uriini eritumist.
Aldosteroon on kõige aktiivsem mineralokortikosteroid, mida neerupealiste koores sünteesitakse kolesteroolist.
Aldosterooni sünteesi ja sekretsiooni glomerulaarse tsooni rakkude poolt stimuleerivad angiotensiin II, ACTH, prostaglandiin E. Need protsessid aktiveeritakse ka kõrge K + ja madala Na + kontsentratsiooni korral.
Hormoon tungib sihtrakku ja interakteerub spetsiifilise retseptoriga, mis asub nii tsütosoolis kui ka tuumas.
Neerutuubulite rakkudes stimuleerib aldosteroon erinevaid funktsioone täitvate valkude sünteesi. Need valgud võivad: a) suurendada naatriumikanalite aktiivsust distaalsete neerutuubulite rakumembraanis, hõlbustades seeläbi naatriumioonide transporti uriinist rakkudesse; b) olla TCA tsükli ensüümid ja seetõttu suurendavad Krebsi tsükli võimet genereerida ioonide aktiivseks transpordiks vajalikke ATP molekule; c) aktiveerida pumba K +, Na + -ATPaasi töö ja stimuleerida uute pumpade sünteesi. Aldosterooni poolt indutseeritud valkude toime üldine tulemus on naatriumioonide reabsorptsiooni suurenemine nefronite tuubulites, mis põhjustab NaCl peetust organismis.
Aldosterooni sünteesi ja sekretsiooni reguleerimise peamine mehhanism on reniin-angiotensiini süsteem.
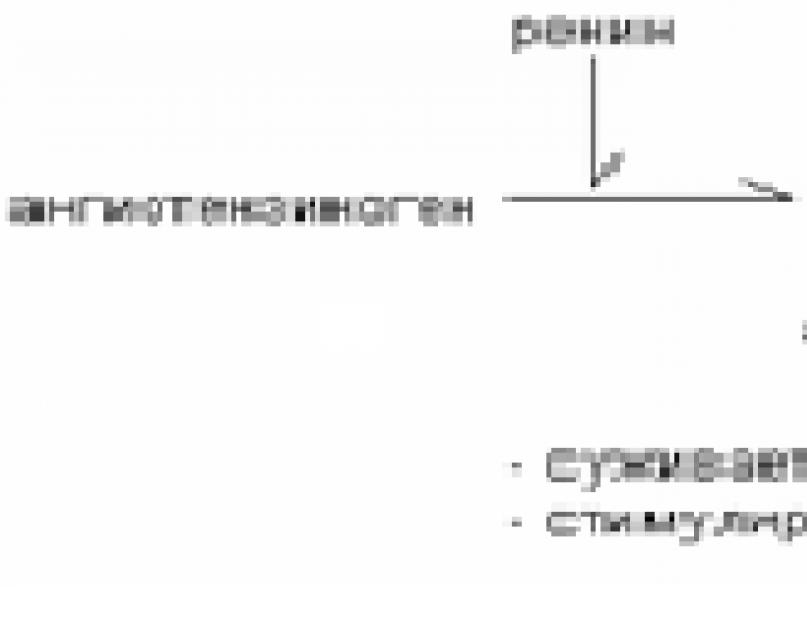
Reniin on ensüüm, mida toodavad neeru aferentsete arterioolide jukstaglomerulaarsed rakud. Nende rakkude lokaliseerimine muudab need vererõhu muutuste suhtes eriti tundlikuks. Vererõhu langus, vedeliku või vere kadu, NaCl kontsentratsiooni langus stimuleerivad reniini vabanemist.
Angiotensinogeen-2 on maksas toodetud globuliin. See toimib reniini substraadina. Reniin hüdrolüüsib peptiidsideme angiotensinogeeni molekulis ja lõikab lahti N-terminaalse dekapeptiidi (angiotensiin I).
Angiotensiin I toimib substraadina antiotensiini konverteeriva ensüümi karboksüdipeptidüülpeptidaasi jaoks, mida leidub endoteelirakkudes ja vereplasmas. Kaks terminaalset aminohapet lõhustatakse angiotensiin I-st, moodustades oktapeptiidi, angiotensiin II.
Angiotensiin II stimuleerib aldosterooni tootmist, põhjustab arterioolide ahenemist, mille tulemuseks on vererõhu tõus ja janu. Angiotensiin II aktiveerib inositoolfosfaadi süsteemi kaudu aldosterooni sünteesi ja sekretsiooni.
PNP on 28 aminohappest koosnev peptiid, millel on üks disulfiidsild. PNP sünteesitakse ja säilitatakse preprohormoonina (koosneb 126 aminohappejäägist) südamerakkudes.
Peamine PNP sekretsiooni reguleeriv tegur on vererõhu tõus. Muud stiimulid: plasma osmolaarsuse suurenemine, südame löögisageduse tõus, katehhoolamiinide ja glükokortikoidide sisalduse tõus veres.
PNP peamised sihtorganid on neerud ja perifeersed arterid.
PNP toimemehhanismil on mitmeid funktsioone. Plasmamembraani PNP retseptor on guanülaattsüklaasi aktiivsusega valk. Retseptoril on domeeni struktuur. Ligandi siduv domeen paikneb ekstratsellulaarses ruumis. PNP puudumisel on PNP retseptori rakusisene domeen fosforüülitud olekus ja inaktiivne. PNP retseptoriga seondumise tulemusena suureneb retseptori guanülaattsüklaasi aktiivsus ja GTP-st moodustub tsükliline GMP. PNP toime tulemusena pärsitakse reniini ja aldosterooni moodustumist ja sekretsiooni. PNP toime üldine mõju on Na + ja vee eritumise suurenemine ning vererõhu langus.
PNP-d peetakse tavaliselt angiotensiin II füsioloogiliseks antagonistiks, kuna selle mõjul ei kaasne veresoonte valendiku ahenemine ja (aldosterooni sekretsiooni reguleerimise kaudu) naatriumi peetus, vaid vastupidi, vasodilatatsioon ja soolade kadu.
Funktsionaalses mõttes on tavaks eristada vaba ja seotud vett. Transpordifunktsioon, mida vesi universaalse lahustina täidab Määrab soolade dissotsiatsiooni dielektrikuna Osalemine erinevates keemilistes reaktsioonides: hüdratsioon hüdrolüüs redoksreaktsioonid näiteks β - rasvhapete oksüdatsioon. Vee liikumine kehas toimub mitmete tegurite osalusel, mille hulka kuuluvad: erineva kontsentratsiooniga soolade tekitatud osmootne rõhk, vesi liigub kõrgema ...
Jagage tööd sotsiaalvõrgustikes
Kui see töö teile ei sobi, on lehe allosas nimekiri sarnastest töödest. Võite kasutada ka otsingunuppu
1. lehekülg
abstraktne
VEE/SOOLA AINEVAHETUS
veevahetus
Vee kogusisaldus täiskasvanud inimese kehas on 60 65% (umbes 40 liitrit). Aju ja neerud on kõige rohkem hüdreeritud. Rasv, luukude, vastupidi, sisaldavad väikeses koguses vett.
Vesi kehas jaotub erinevates osakondades (sektsioonides, basseinides): rakkudes, rakkudevahelises ruumis, anumate sees.
Intratsellulaarse vedeliku keemilise koostise tunnuseks on kõrge kaaliumi- ja valkude sisaldus. Ekstratsellulaarne vedelik sisaldab suuremas kontsentratsioonis naatriumi. Ekstratsellulaarse ja rakusisese vedeliku pH väärtused ei erine. Funktsionaalses mõttes on tavaks eristada vaba ja seotud vett. Seotud vesi on see osa sellest, mis on osa biopolümeeride hüdratatsioonikestast. Seotud vee hulk iseloomustab intensiivsust metaboolsed protsessid.
Vee bioloogiline roll organismis.
- Transpordifunktsioon, mida vesi universaalse lahustina täidab
- Määrab soolade dissotsiatsiooni, olles dielektrik
- Osalemine erinevates keemilistes reaktsioonides: hüdratsioon, hüdrolüüs, redoksreaktsioonid (näiteks β - rasvhapete oksüdatsioon).
Veevahetus.
Täiskasvanu vahetatava vedeliku kogumaht on 2-2,5 liitrit päevas. Täiskasvanu iseloomustab veetasakaal, s.t. vedeliku tarbimine on võrdne selle eritumisega.
Vesi siseneb kehasse vedelate jookide kujul (umbes 50% tarbitavast vedelikust), tahke toidu osana. 500 ml on endogeenne vesi, mis moodustub kudedes oksüdatiivsete protsesside tulemusena,
Vee eritumine organismist toimub neerude kaudu (1,5 l diurees), aurustumisel naha pinnalt, kopsudest (umbes 1 l), soolte kaudu (umbes 100 ml).
Vee liikumise tegurid kehas.
Vesi kehas jaotub pidevalt ümber erinevate sektsioonide vahel. Vee liikumine kehas toimub mitmete tegurite osalusel, sealhulgas:
- osmootne rõhk, mille tekitavad erinevad soolakontsentratsioonid (vesi liigub suurema soolakontsentratsiooni poole),
- onkootiline rõhk, mis tekib valgu kontsentratsiooni langusest (vesi liigub kõrgema valgukontsentratsiooni suunas)
- südame poolt tekitatud hüdrostaatiline rõhk
Veevahetus on vahetusega tihedalt seotud Na ja K.
Naatriumi ja kaaliumi vahetus
Kindral naatriumi sisalduskehas on 100 g Samal ajal langeb 50% ekstratsellulaarsele naatriumile, 45% - luudes sisalduvale naatriumile, 5% - intratsellulaarsele naatriumile. Naatriumisisaldus vereplasmas on 130-150 mmol/l, vererakkudes - 4-10 mmol/l. Täiskasvanu naatriumivajadus on umbes 4-6 g päevas.
Kindral kaaliumisisaldustäiskasvanud inimese kehas on 160 90% sellest kogusest sisaldub intratsellulaarselt, 10% jaotub rakuvälises ruumis. Vereplasma sisaldab 4-5 mmol / l, rakkude sees - 110 mmol / l. Täiskasvanu päevane kaaliumivajadus on 2-4 g.
Naatriumi ja kaaliumi bioloogiline roll:
- Osmootse rõhu määramine
- määrata vee jaotus
- luua vererõhku
- osalema (na ) aminohapete, monosahhariidide imendumisel
- kaalium on biosünteesiprotsesside jaoks hädavajalik.
Naatriumi ja kaaliumi imendumine toimub maos ja sooltes. Naatrium võib maksas veidi ladestuda. Naatrium ja kaalium erituvad organismist peamiselt neerude, vähesel määral higinäärmete ja soolte kaudu.
Osaleb naatriumi ja kaaliumi ümberjaotamises rakkude ja rakuvälise vedeliku vahelnaatrium - kaalium ATPaas -membraaniensüüm, mis kasutab ATP energiat naatriumi- ja kaaliumiioonide liigutamiseks kontsentratsioonigradienti vastu. Tekkinud erinevus naatriumi ja kaaliumi kontsentratsioonis tagab kudede ergastamise protsessi.
Vee-soola ainevahetuse reguleerimine.
Vee ja soolade vahetuse reguleerimine toimub kesknärvisüsteemi, autonoomse närvisüsteemi ja endokriinsüsteemi osalusel.
Kesknärvisüsteemis tekib vedeliku hulga vähenemisega kehas janutunne. Hüpotalamuses asuva joogikeskuse ergastamine viib vee tarbimiseni ja selle koguse taastamiseni organismis.
Autonoomne närvisüsteem osaleb vee ainevahetuse reguleerimises, reguleerides higistamisprotsessi.
Vee ja soolade ainevahetuse reguleerimises osalevate hormoonide hulka kuuluvad antidiureetiline hormoon, mineralokortikoidid, natriureetiline hormoon.
Antidiureetiline hormoonsünteesitakse hüpotalamuses, liigub hüpofüüsi tagumisse osasse, kust vabaneb verre. See hormoon säilitab vett kehas, suurendades vee vastupidist tagasiimendumist neerudes, aktiveerides neis akvaporiini valgu sünteesi.
Aldosteroon aitab kaasa naatriumi säilimisele organismis ja kaaliumiioonide kadumisele neerude kaudu. Arvatakse, et see hormoon soodustab naatriumikanali valkude sünteesi, mis määravad naatriumi vastupidise reabsorptsiooni. Samuti aktiveerib see Krebsi tsükli ja ATP sünteesi, mis on vajalik naatriumi reabsorptsiooni protsessideks. Aldosteroon aktiveerib valkude – kaaliumi transporterite sünteesi, millega kaasneb kaaliumi suurenenud eritumine organismist.
Nii antidiureetilise hormooni kui ka aldosterooni funktsioon on tihedalt seotud vere reniini-angiotensiini süsteemiga.
Reniini-angiotensiivne veresüsteem.
Neerude verevoolu vähenemisega dehüdratsiooni ajal toodetakse neerudes proteolüütilist ensüümi reniin, mis tõlgibangiotensinogeen(α2-globuliin) angiotensiin I-ks - 10 aminohappest koosnev peptiid. Angiotensiin Olen tegevuse all angiotesiini konverteeriv ensüüm(ACE) läbib täiendava proteolüüsi ja läheb edasi angiotensiin II , sealhulgas 8 aminohapet, angiotensiin II ahendab veresooni, stimuleerib antidiureetilise hormooni ja aldosterooni tootmist, mis suurendavad vedeliku mahtu kehas.

Natriureetiline peptiidtoodetakse kodades vastusena vee mahu suurenemisele kehas ja kodade venitamisele. See koosneb 28 aminohappest, on disulfiidsildadega tsükliline peptiid. Natriureetiline peptiid soodustab naatriumi ja vee väljutamist organismist.
Vee-soola ainevahetuse rikkumine.
Vee ja soolade ainevahetuse häired hõlmavad dehüdratsiooni, hüperhüdratsiooni, naatriumi ja kaaliumi kontsentratsiooni kõrvalekaldeid vereplasmas.
Dehüdratsioon (dehüdratsiooniga) kaasneb tõsine kesknärvisüsteemi talitlushäire. Dehüdratsiooni põhjused võivad olla:
- veenälg,
- soole düsfunktsioon (kõhulahtisus),
- suurenenud kaotus kopsude kaudu (õhupuudus, hüpertermia),
- suurenenud higistamine,
- suhkur ja mitte diabeet.
Hüperhüdratsioonveekoguse suurenemist kehas võib täheldada mitmete patoloogiliste seisundite korral:
- suurenenud vedeliku tarbimine kehas,
- neerupuudulikkus,
- vereringehäired,
- maksahaigus
Kohalik ilming vedeliku kogunemine kehas on turse.
"Näljast" turset täheldatakse hüpoproteineemia tõttu valgu nälgimise ajal, maksahaigused. "Südame" turse tekib siis, kui südamehaiguste korral on häiritud hüdrostaatiline rõhk. "Neeruturse" tekib siis, kui neeruhaiguste korral muutub vereplasma osmootne ja onkootiline rõhk
Hüponatreemia, hüpokaleemiaavalduvad erutuvuse rikkumises, närvisüsteemi kahjustuses, südamerütmi rikkumises. Need seisundid võivad ilmneda mitmesuguste patoloogiliste seisundite korral:
- neerufunktsiooni häired
- korduv oksendamine
- kõhulahtisus
- aldosterooni, natriureetilise hormooni tootmise rikkumine.
Neerude roll vee-soola ainevahetuses.
Neerudes toimub filtreerimine, reabsorptsioon, naatriumi, kaaliumi sekretsioon. Neere reguleerib aldosteroon, antidiureetiline hormoon. Neerud toodavad reniini, reniini lähteensüümi, angiotensiini süsteemi. Neerud eritavad prootoneid ja reguleerivad seeläbi pH-d.
Vee ainevahetuse tunnused lastel.
Lastel suureneb kogu veesisaldus, mis vastsündinutel ulatub 75% -ni. Lapsepõlves täheldatakse erinevat vee jaotumist kehas: rakusisese vee kogus väheneb 30% -ni, mis on tingitud rakusiseste valkude sisalduse vähenemisest. Samal ajal suurendati rakuvälise vee sisaldust kuni 45%, mis on seotud hüdrofiilsete glükoosaminoglükaanide suurema sisaldusega sidekoe rakkudevahelises aines.
Vee ainevahetus lapse kehas toimub intensiivsemalt. Lastel on veevajadus 2-3 korda suurem kui täiskasvanutel. Lastele on iseloomulik see, et seedemahlas vabaneb suur kogus vett, mis imendub kiiresti tagasi. Väikelastel erinev veekao suhe organismist: suurem osa veest väljub kopsude ja naha kaudu. Lastele on iseloomulik veepeetus kehas (positiivne veetasakaal)
Lapsepõlves täheldatakse vee ainevahetuse ebastabiilset regulatsiooni, janutunnet ei teki, mille tagajärjel väljendub kalduvus dehüdratsioonile.
Esimestel eluaastatel on kaaliumi eritumine ülekaalus naatriumi eritumisest.
Kaltsium-fosfori metabolism
Üldine sisu kaltsium on 2% kehakaalust (umbes 1,5 kg). 99% sellest on koondunud luudesse, 1% on rakuväline kaltsium. Kaltsiumisisaldus vereplasmas on võrdne 2,3-2,8 mmol/l, 50% sellest kogusest on ioniseeritud kaltsium ja 50% valkudega seotud kaltsium.
Kaltsiumi funktsioonid:
- plastmaterjal
- osaleb lihaste kontraktsioonis
- osalevad vere hüübimises
- paljude ensüümide aktiivsuse regulaator (mängib teise sõnumitooja rolli)
Täiskasvanu päevane kaltsiumivajadus on 1,5 g Kaltsiumi imendumine seedetraktis on piiratud. Osalemisel imendub ligikaudu 50% toiduga saadavast kaltsiumistkaltsiumi siduv valk. Olles ekstratsellulaarne katioon, siseneb kaltsium rakkudesse kaltsiumikanalite kaudu, ladestub rakkudesse sarkoplasmaatilises retikulumis ja mitokondrites.
Üldine sisu fosforit kehas on 1% kehakaalust (umbes 700 g). 90% fosforist leidub luudes, 10% on rakusisene fosfor. Vereplasmas on fosforisisaldus 1 -2 mmol/l
Fosfori funktsioonid:
- plastiline funktsioon
- on osa makroergsist (ATP)
- nukleiinhapete, lipoproteiinide, nukleotiidide, soolade komponent
- osa fosfaatpuhvrist
- paljude ensüümide aktiivsuse regulaator (ensüümide fosforüülimine defosforüülimine)
Täiskasvanu päevane fosforivajadus on umbes 1,5 g.Seedetraktis imendub fosfor osaluselaluseline fosfataas.
Kaltsium ja fosfor erituvad organismist peamiselt neerude kaudu, väike kogus läheb kaduma soolte kaudu.
Kaltsiumfosfori metabolismi reguleerimine.
Kaltsiumi ja fosfori ainevahetuse reguleerimises osalevad paratüroidhormoon, kaltsitoniin, D-vitamiin.
Parathormoon suurendab kaltsiumi taset veres ja samal ajal vähendab fosfori taset. Kaltsiumisisalduse suurenemine on seotud aktiveerimisegafosfataasid, kollagenaasidosteoklastid, mille tulemusena luukoe uuenemisel "pestakse" kaltsiumi verre. Lisaks aktiveerib paratüreoidhormoon kaltsiumi siduva valgu osalusel kaltsiumi imendumist seedetraktis ja vähendab kaltsiumi eritumist neerude kaudu. Paratüreoidhormooni toimel olevad fosfaadid, vastupidi, erituvad intensiivselt neerude kaudu.
Kaltsitoniin vähendab kaltsiumi ja fosfori taset veres. Kaltsitoniin vähendab osteoklastide aktiivsust ja vähendab seeläbi kaltsiumi vabanemist luukoest.
D-vitamiin kolekaltsiferool, anti-rahhiitne vitamiin.
D-vitamiin viitab rasvlahustuvatele vitamiinidele. Päevane vitamiinivajadus on 25 mcg. D-vitamiin UV-kiirte mõjul sünteesitakse see nahas oma eelkäijast 7-dehüdrokolesteroolist, mis koos valguga siseneb maksa. Maksas toimub oksügenaaside mikrosomaalse süsteemi osalusel oksüdatsioon 25. positsioonil koos 25-hüdroksükolekaltsiferooli moodustumisega. See vitamiini prekursor kantakse spetsiifilise transpordivalgu osalusel neerudesse, kus see läbib teise hüdroksüülimisreaktsiooni esimeses positsioonis koos moodustumisega. D3-vitamiini aktiivne vorm - 1,25-dihüdrokolekaltsiferool (või kaltsitriool). . Hüdroksüülimisreaktsiooni neerudes aktiveerib paratüreoidhormoon, kui kaltsiumi tase veres väheneb. Piisava kaltsiumisisaldusega organismis moodustub neerudes inaktiivne metaboliit 24,25 (OH). C-vitamiin osaleb hüdroksüülimisreaktsioonides.
1,25 (OH) 2 D 3 toimib sarnaselt steroidhormoonidega. Tungides sihtrakkudesse, interakteerub see retseptoritega, mis migreeruvad rakutuuma. Enterotsüütides stimuleerib see hormooni retseptori kompleks valgu kaltsiumi kandja sünteesi eest vastutava mRNA transkriptsiooni. Soolestikus paraneb kaltsiumi imendumine kaltsiumi siduva valgu ja Ca osalusel 2+ - ATPaasid. Luukoes vitamiin D3 stimuleerib demineralisatsiooni protsessi. Neerudes aktiveerumine vitamiiniga D3 kaltsiumi ATP-aasiga kaasneb kaltsiumi- ja fosfaadiioonide reabsorptsiooni suurenemine. Kaltsitriool osaleb luuüdi rakkude kasvu ja diferentseerumise reguleerimises. Sellel on antioksüdantne ja kasvajavastane toime.
Hüpovitaminoos põhjustab rahhiidi.
Hüpervitaminoos põhjustab luude tugevat demineraliseerumist, pehmete kudede lupjumist.
Kaltsiumfosfori metabolismi rikkumine
Rahhiit mis väljendub luukoe mineraliseerumise häiretes. Haiguse põhjuseks võib olla hüpovitaminoos D3. , päikesevalguse puudumine, keha ebapiisav tundlikkus vitamiini suhtes. Rahhiidi biokeemilisteks sümptomiteks on kaltsiumi ja fosfori taseme langus veres ning aluselise fosfataasi aktiivsuse vähenemine. Lastel avaldub rahhiit osteogeneesi rikkumises, luude deformatsioonides, lihaste hüpotensioonis ja suurenenud neuromuskulaarses erutuvuses. Täiskasvanutel põhjustab hüpovitaminoos kaariese ja osteomalaatsia, eakatel - osteoporoosi.
Vastsündinutel võib arenedamööduv hüpokaltseemia, kuna kaltsiumi omastamine ema kehast peatub ja täheldatakse hüpoparatüreoidismi.
Hüpokaltseemia, hüpofosfateemiavõib tekkida paratüreoidhormooni, kaltsitoniini tootmise, seedetrakti düsfunktsiooni (oksendamine, kõhulahtisus), neerude, obstruktiivse ikterusega, luumurdude paranemise ajal.
Rauavahetus.
Üldine sisu nääre täiskasvanud inimese organismis on 5 g Raud jaotub peamiselt rakusiseselt, kus on ülekaalus heemraud: hemoglobiin, müoglobiin, tsütokroomid. Ekstratsellulaarset rauda esindab valk transferriin. Vereplasmas on rauasisaldus 16-19 µmol/l, erütrotsüütides - 19 mmol/l. O Raua ainevahetus täiskasvanutel on 20-25 mg päevas . Põhiosa sellest kogusest (90%) moodustab endogeenne raud, mis vabaneb erütrotsüütide lagunemisel, 10% on eksogeenne raud, mida tarnitakse toiduainete osana.
Raua bioloogilised funktsioonid:
- organismi redoksprotsesside oluline komponent
- hapniku transport (hemoglobiini osana)
- hapniku ladestumine (müoglobiini koostises)
- antioksüdantne funktsioon (katalaasi ja peroksüdaaside osana)
- stimuleerib immuunvastuseid organismis
Raua imendumine toimub soolestikus ja see on piiratud protsess. Arvatakse, et 1/10 toidus leiduvast rauast imendub. Toiduained sisaldavad oksüdeeritud 3-valentset rauda, mis mao happelises keskkonnas muutub F e 2+ . Raua imendumine toimub mitmel etapil: sisenemine enterotsüütidesse limaskesta mutsiini osalusel, intratsellulaarne transport enterotsüütide ensüümide abil ja raua üleminek vereplasmasse. Valk, mis osaleb raua imendumises apoferritiin, mis seob rauda ja jääb soole limaskestale, luues raua depoo. See raua metabolismi etapp on regulatiivne: apoferritiini süntees väheneb raua puudumisega organismis.
Imendunud raud transporditakse transferriini valgu osana, kus see oksüdeerubtseruloplasmiin kuni F e 3+ , mille tulemusena suureneb raua lahustuvus. Transferriin interakteerub kudede retseptoritega, mille arv on väga erinev. See vahetuse etapp on samuti regulatiivne.
Raud võib ladestuda ferritiini ja hemosideriini kujul. ferritiin maksa vees lahustuv valk, mis sisaldab kuni 20% F e 2+ fosfaadi või hüdroksiidina. Hemosideriin lahustumatu valk, sisaldab kuni 30% F e 3+ , sisaldab oma koostises polüsahhariide, nukleotiide, lipiide ..
Raua eritumine organismist toimub naha ja soolte kooriva epiteeli osana. Väike kogus rauda kaob neerude kaudu sapi ja süljega.
Raua metabolismi kõige levinum patoloogia onRauavaegusaneemia.Samas on hemosideriini kuhjumise ja arenguga võimalik ka keha rauaga üleküllastada. hemokromatoos.
KUDE BIOKEEMIA
Sidekoe biokeemia.
Erinevat tüüpi sidekude on ehitatud ühe põhimõtte järgi: kiud (kollageen, elastiin, retikuliin) ja erinevad rakud (makrofaagid, fibroblastid ja muud rakud) on jaotunud suures massis rakkudevahelises põhiaines (proteoglükaanid ja retikulaarsed glükoproteiinid).
Sidekude täidab mitmeid funktsioone:
- tugifunktsioon (luu skelett),
- barjäärifunktsioon
- metaboolne funktsioon (koe keemiliste komponentide süntees fibroblastides),
- ladestusfunktsioon (melaniini akumuleerumine melanotsüütides),
- reparatiivne funktsioon (osalemine haavade paranemises),
- osalemine vee-soola ainevahetuses (proteoglükaanid seovad rakuvälist vett)
Peamise rakkudevahelise aine koostis ja vahetus.
Proteoglükaanid (vt süsivesikute keemia) ja glükoproteiinid (ibid.).
Glükoproteiinide ja proteoglükaanide süntees.
Proteoglükaanide süsivesikute komponenti esindavad glükoosaminoglükaanid (GAG), mille hulka kuuluvad atsetüülaminosuhkrud ja uroonhapped. Nende sünteesi lähteaineks on glükoos.
- glükoos-6-fosfaat → fruktoos-6-fosfaat glutamiin → glükoosamiin.
- glükoos → UDP-glükoos →UDP - glükuroonhape
- glükoosamiin + UDP-glükuroonhape + FAPS → GAG
- GAG + valk → proteoglükaan
proteoglükaanide ja glükoproteiinide laguneminemida teostavad erinevad ensüümid: hüaluronidaas, iduronidaas, heksaminidaas, sulfataas.
Sidekoe valkude metabolism.
Kollageenivahetus
Sidekoe peamine valk on kollageen (vt struktuuri jaotises "Valkude keemia"). Kollageen on polümorfne valk, mille koostises on mitmesugused polüpeptiidahelate kombinatsioonid. Inimkehas domineerivad 1,2,3 tüüpi kollageeni fibrillid moodustavad vormid.
Kollageeni süntees.
Kollageeni süntees toimub firoblastides ja rakuvälises ruumis, hõlmab mitmeid etappe. Esimestel etappidel sünteesitakse prokollageeni (esindatud 3 polüpeptiidahelaga, millel on täiendavad N ja C-otsa fragmendid). Seejärel toimub prokollageeni translatsioonijärgne modifikatsioon kahel viisil: oksüdatsiooni (hüdroksüülimise) ja glükosüülimise teel.
- aminohapped lüsiin ja proliin läbivad ensüümide osalusel oksüdatsioonilüsiini oksügenaas, proliinoksügenaas, raua ioonid ja C-vitamiin.Saadud hüdroksülüsiin, hüdroksüproliin, osaleb kollageenis ristsidemete moodustamises
- süsivesikute komponendi kinnitamine toimub ensüümide osaluselglükosüültransferaasid.
Modifitseeritud prokollageen siseneb rakkudevahelisse ruumi, kus see läbib osalise proteolüüsi terminali lõhustamise teel N ja C fragmendid. Selle tulemusena muundatakse prokollageen tropokollageen - kollageenkiudude struktuurne plokk.
Kollageeni lagunemine.
Kollageen on aeglaselt vahetuv valk. Kollageeni lagundamine toimub ensüümi toimel kollagenaas. See on tsinki sisaldav ensüüm, mis sünteesitakse prokollagenaasina. Prokollagenaas aktiveeritaksetrüpsiin, plasmiin, kallikreiinosalise proteolüüsi teel. Kollagenaas lagundab molekuli keskel asuva kollageeni suurteks fragmentideks, mida tsinki sisaldavad ensüümid täiendavalt lõhustavad.želatinaasid.
C-vitamiin, askorbiinhape, antiskorbüütiline vitamiin
C-vitamiin mängib kollageeni metabolismis väga olulist rolli. Keemiliselt on see laktoonhape, mis on oma struktuurilt sarnane glükoosiga. Täiskasvanu päevane askorbiinhappe vajadus on 50 100 mg. C-vitamiini leidub puu- ja köögiviljades. C-vitamiini roll on järgmine:
- osaleb kollageeni sünteesis,
- osaleb türosiini metabolismis,
- osaleb foolhappe üleminekul THFA-ks,
- on antioksüdant
Avitaminoos "C" avaldub skorbuut (gingiviit, aneemia, verejooks).
Elastiini vahetus.
Elastiini vahetust ei mõisteta hästi. Arvatakse, et elastiini süntees proelastiini kujul toimub ainult embrüo perioodil. Elastiini lagundamine toimub neutrofiilide ensüümi toimel elastaas , mis sünteesitakse mitteaktiivse proelastaasina.
Sidekoe koostise ja ainevahetuse tunnused lapsepõlves.
- Suurem proteoglükaanide sisaldus,
- GAG-ide erinev suhe: rohkem hüaluroonhapet, vähem kondrotiinsulfaate ja kerataansulfaate.
- Domineerib 3. tüüpi kollageen, mis on vähem stabiilne ja vahetub kiiremini.
- Sidekoe komponentide intensiivsem vahetus.
Sidekoe häired.
Glükoosaminoglükaanide ja proteoglükaanide metabolismi võimalikud kaasasündinud häiredmukopolüsahharidoosid.Teine sidekoehaiguste rühm on kollagenoos, eriti reuma. Kollagenooside korral täheldatakse kollageeni hävimist, mille üheks sümptomiks onhüdroksüprolinuuria
Vöötlihaskoe biokeemia
Lihaste keemiline koostis: 80-82% on vesi, 20% kuivjääk. 18% kuivjäägist langeb valkudele, ülejäänud osa moodustavad lämmastikku sisaldavad mittevalgulised ained, lipiidid, süsivesikud ja mineraalid.
Lihasvalgud.
Lihasvalgud jagunevad kolme tüüpi:
- sarkoplasmaatilised (vees lahustuvad) valgud moodustavad 30% kõigist lihasvalkudest
- müofibrillaarsed (soolalahustuvad) valgud moodustavad 50% kõigist lihasvalkudest
- strooma (vees lahustumatud) valgud moodustavad 20% kõigist lihasvalkudest
Müofibrillaarsed valgudmida esindavad müosiin, aktiin, (peamised valgud) tropomüosiin ja troponiin (väikesed valgud).
müosiin - Müofibrillide paksude filamentide valk, mille molekulmass on umbes 500 000 päeva, koosneb kahest raskest ahelast ja 4 kergest ahelast. Müosiin kuulub kera-fibrillaarsete valkude rühma. See vaheldub kergete ahelate kerakujulisi "päid" ja raskete ahelate fibrillaarseid "sabasid". Müosiini "pea" on ensümaatilise ATPaasi aktiivsusega. Müosiin moodustab 50% müofibrillaarsetest valkudest.
aktiin esitatud kahel kujul kerakujuline (G-vorm), fibrillaarne (F-vorm). G-kujuline selle molekulmass on 43 000 d. F -aktiini vorm on sfääriliste keerdunud filamentide kujul G -vormid. See valk moodustab 20-30% müofibrillaarsetest valkudest.
Tropomüosiin - väike valk molekulmassiga 65 000 g. See on ovaalse varda kujuga, sobib aktiivhõõgniidi süvenditesse ning täidab "isolaatori" funktsiooni aktiiv- ja müosiini filamendi vahel.
Troponiin Ca on sõltuv valk, mis muudab kaltsiumiioonidega suhtlemisel oma struktuuri.
Sarkoplasmaatilised valgudmida esindavad müoglobiin, ensüümid, hingamisahela komponendid.
Strooma valgud - kollageen, elastiin.
Lihaste lämmastikku sisaldavad ekstraktsiooniained.
Lämmastikku sisaldavate mittevalguliste ainete hulka kuuluvad nukleotiidid (ATP), aminohapped (eriti glutamaat), lihasedipeptiidid (karnosiin ja anseriin). Need dipeptiidid mõjutavad naatriumi- ja kaltsiumipumpade tööd, aktiveerivad lihaste tööd, reguleerivad apoptoosi ja on antioksüdandid. Lämmastikku sisaldavate ainete hulka kuuluvad kreatiin, fosfokreatiin ja kreatiniin. Kreatiin sünteesitakse maksas ja transporditakse lihastesse.
Orgaanilised lämmastikuvabad ained
Lihased sisaldavad kõiki klasse lipiidid. Süsivesikud mida esindavad glükoos, glükogeen ja süsivesikute ainevahetuse saadused (laktaat, püruvaat).
Mineraalid
Lihased sisaldavad palju mineraale. Suurim kaltsiumi, naatriumi, kaaliumi, fosfori kontsentratsioon.
Lihaste kokkutõmbumise ja lõõgastumise keemia.
Kui vöötlihased on erutatud, vabanevad kaltsiumiioonid sarkoplasmaatilisest retikulumist tsütoplasmasse, kus Ca kontsentratsioon 2+ suureneb 10-ni-3 palvetama. Kaltsiumiioonid interakteeruvad regulatoorse valgu troponiiniga, muutes selle konformatsiooni. Selle tulemusena nihkub reguleeriv valk tropomüosiin mööda aktiinikiudu ning aktiini ja müosiini vahelised interaktsioonikohad vabanevad. Müosiini ATPaasi aktiivsus aktiveerub. ATP energia tõttu muutub müosiini "pea" kaldenurk "saba" suhtes ja selle tulemusena libisevad aktiini filamendid müosiini filamentide suhtes.lihaste kokkutõmbumine.
Impulsside lõppemisel "pumbatakse" kaltsiumiioonid ATP energia tõttu Ca-ATP-aasi osalusel sarkoplasmaatilisesse retikulumi. Ca kontsentratsioon 2+ tsütoplasmas väheneb 10-ni-7 mool, mis viib troponiini vabanemiseni kaltsiumiioonidest. Sellega omakorda kaasneb kontraktiilsete valkude aktiini ja müosiini eraldamine tropomüosiini valgu poolt. lihaste lõdvestamine.
Lihaste kokkutõmbumiseks kasutatakse järjestikku järgmist:energiaallikad:
- piiratud endogeense ATP pakkumine
- ebaoluline kreatiinfosfaadi fond
- ATP moodustumine 2 ADP molekuli tõttu ensüümi müokinaasi osalusel
(2 ADP → AMP + ATP)
- anaeroobne glükoosi oksüdatsioon
- glükoosi, rasvhapete, atsetooni kehade oksüdatsiooni aeroobsed protsessid
Lapsepõlvesveesisaldus lihastes on suurenenud, müofibrillaarsete valkude osakaal on väiksem, stroomavalkude tase kõrgem.
Vöötlihaste keemilise koostise ja funktsiooni rikkumised hõlmavad müopaatia, mille puhul on lihaste energiavahetuse rikkumine ja müofibrillaarsete kontraktiilsete valkude sisalduse vähenemine.
Närvikoe biokeemia.
Aju hallaine (neuronikehad) ja valgeaine (aksonid) erinevad vee ja lipiidide sisalduse poolest. Halli ja valge aine keemiline koostis:

aju valgud
aju valguderinevad lahustuvuse poolest. Eraldadavees lahustuv(soollahustuvad) närvikoe valgud, mille hulka kuuluvad neuroalbumiinid, neuroglobuliinid, histoonid, nukleoproteiinid, fosfoproteiinid javees lahustumatu(soolas lahustumatu), mille hulka kuuluvad neurokollageen, neuroelastiin, neurostromiin.
Lämmastikku sisaldavad mittevalgulised ained
Aju mittevalgulisi lämmastikku sisaldavaid aineid esindavad aminohapped, puriinid, kusihape, karnosiindipeptiid, neuropeptiidid, neurotransmitterid. Aminohapetest leidub suuremas kontsentratsioonis glutamaati ja aspatraati, mis on seotud aju ergastavate aminohapetega.
Neuropeptiidid (neuroenkefaliinid, neuroendorfiinid) need on peptiidid, millel on morfiinitaoline valuvaigistav toime. Need on immunomodulaatorid, täidavad neurotransmitteri funktsiooni. neurotransmitterid norepinefriin ja atsetüülkoliin on biogeensed amiinid.
Aju lipiidid
Lipiidid moodustavad halli massist 5% ja valgeaine märgmassist 17%, aju kuivmassist vastavalt 30–70%. Närvikoe lipiide esindavad:
- vabad rasvhapped (arahhidoon-, tserebroon-, närvi-)
- fosfolipiidid (atsetalfosfatiidid, sfingomüeliinid, koliinfosfatiidid, kolesterool)
- sfingolipiidid (gangliosiidid, tserebrosiidid)
Rasvade jaotus hallis ja valges aines on ebaühtlane. Hallis on madalam kolesteroolisisaldus, kõrge tserebrosiidide sisaldus. Valgeaines on kolesterooli ja gangliosiidide osakaal suurem.
aju süsivesikud
Süsivesikud sisalduvad ajukoes väga madalates kontsentratsioonides, mis on tingitud glükoosi aktiivsest kasutamisest närvikoes. Süsivesikuid esindab glükoos kontsentratsioonis 0,05%, süsivesikute metabolismi metaboliidid.
Mineraalid
Naatrium, kaltsium, magneesium jaotuvad hallis ja valges aines üsna ühtlaselt. Valgeaines on suurenenud fosfori kontsentratsioon.
Närvikoe põhiülesanne on närviimpulsside juhtimine ja edastamine.
Närviimpulsi juhtimine
Närviimpulsi juhtivus on seotud naatriumi ja kaaliumi kontsentratsiooni muutumisega rakkude sees ja väljaspool. Kui närvikiud on ergastatud, suureneb järsult neuronite ja nende protsesside läbilaskvus naatriumile. Naatrium rakuvälisest ruumist siseneb rakkudesse. Kaaliumi vabanemine rakkudest viibib. Selle tulemusena tekib membraanile laeng: välispind omandab negatiivse ja sisepind positiivse laengu.tegevuspotentsiaal. Ergastuse lõpus "pumbatakse" naatriumiioonid K osalusel ekstratsellulaarsesse ruumi, Na -ATPaasi ja membraan laetakse uuesti. Väljaspool on positiivne laeng ja sees - negatiivne laeng - on puhkepotentsiaal.
Närviimpulsi edastamine
Närviimpulsi ülekanne sünapsis toimub sünapsides neurotransmitterite abil. Klassikalised neurotransmitterid on atsetüülkoliin ja norepinefriin.
Atsetüülkoliin sünteesitakse atsetüül-CoA-st ja koliinist ensüümi osaluselatsetüülkoliini transferaas, koguneb sünaptilistesse vesiikulitesse, vabaneb sünaptilisse pilusse ja interakteerub postsünaptilise membraani retseptoritega. Atsetüülkoliini lagundab ensüüm koliinesteraas.
Norepinefriin sünteesitakse türosiinist, mille ensüüm hävitabmonoamiini oksüdaas.


GABA (gamma-aminovõihape), serotoniin ja glütsiin võivad samuti toimida vahendajatena.
Närvikoe ainevahetuse tunnusedon järgmised:
- hematoentsefaalbarjääri olemasolu piirab aju läbilaskvust paljudele ainetele,
- domineerivad aeroobsed protsessid
- Glükoos on peamine energiaallikas
Lastel sünnihetkeks on moodustunud 2/3 neuronitest, ülejäänud moodustuvad esimese aasta jooksul. Üheaastase lapse aju mass moodustab umbes 80% täiskasvanu aju massist. Aju küpsemise protsessis suureneb lipiidide sisaldus järsult ja müeliniseerumisprotsessid toimuvad aktiivselt.
Maksa biokeemia.
Maksakoe keemiline koostis: 80% vesi, 20% kuivjääk (valgud, lämmastikku sisaldavad ained, lipiidid, süsivesikud, mineraalid).
Maks osaleb inimkeha igat tüüpi ainevahetuses.
süsivesikute ainevahetus
Maksas toimub aktiivselt glükogeeni süntees ja lagunemine, glükoneogenees, toimub galaktoosi ja fruktoosi assimilatsioon ning pentoosfosfaadi rada on aktiivne.
lipiidide metabolism
Maksas triatsüülglütseroolide, fosfolipiidide, kolesterooli süntees, lipoproteiinide (VLDL, HDL) süntees, sapphapete süntees kolesteroolist, atsetoonikehade süntees, mis seejärel transporditakse kudedesse,
lämmastiku metabolism
Maksa iseloomustab aktiivne valkude metabolism. See sünteesib kõiki albumiine ja enamikku vereplasma globuliine, vere hüübimisfaktoreid. Maksas tekib ka teatud kehavalkude reserv. Maksas toimub aktiivselt aminohapete katabolism - deamineerimine, transamiinimine, uurea süntees. Hepatotsüütides puriinid lagunevad kusihappe moodustumisega, lämmastikku sisaldavate ainete - koliini, kreatiini sünteesiga.
Antitoksiline funktsioon
Maks on kõige olulisem organ nii eksogeensete (ravimid) kui ka endogeensete toksiliste ainete (bilirubiin, valkude lagunemissaadused, ammoniaak) neutraliseerimiseks. Toksiliste ainete detoksikatsioon maksas toimub mitmel etapil:
- suurendab neutraliseeritud ainete polaarsust ja hüdrofiilsust oksüdatsioon (indool indoksüüliks), hüdrolüüs (atsetüülsalitsüül → äädikhape + salitsüülhape), redutseerimine jne.
- konjugatsioon glükuroonhappe, väävelhappe, glükokooli, glutatiooni, metallotioneiiniga (raskmetallide soolade jaoks)
Biotransformatsiooni tulemusena väheneb toksilisus reeglina märkimisväärselt.
pigmendivahetus
Maksa osalemine sapipigmentide metabolismis seisneb bilirubiini neutraliseerimises, urobilinogeeni hävitamises.
Porfüriini vahetus:
Maks sünteesib porfobilinogeeni, uroporfürinogeeni, koproporfürinogeeni, protoporfüriini ja heemi.
Hormoonide vahetus
Maks inaktiveerib aktiivselt adrenaliini, steroide (konjugatsioon, oksüdatsioon), serotoniini ja teisi biogeenseid amiine.
Maks osaleb kaudselt vee-soola metabolismis, sünteesides vereplasma valke, mis määravad onkootilist rõhku, angiotensinogeeni, angiotensiini prekursori sünteesi. II.
Mineraalide vahetus
: Maksas raua, vase ladestumine, transportvalkude tseruloplasmiini ja transferriini süntees, mineraalide eritumine sapiga.
Varastel lapsepõlvesmaksafunktsioonid on arengujärgus, nende rikkumine on võimalik.
Kirjandus
Barker R.: Demonstratiivne neuroteadus. - M.: GEOTAR-Media, 2005
I.P. Ashmarin, E.P. Karazeeva, M.A. Karabasova ja teised: patoloogiline füsioloogia ja biokeemia. - M.: Eksam, 2005
Kvetnaya T.V.: Melatoniin on vanusega seotud patoloogia neuroimmunoendokriinne marker. - Peterburi: DEAN, 2005
Pavlov A.N.: Ökoloogia: ratsionaalne keskkonnajuhtimine ja eluohutus. - M.: Kõrgkool, 2005
Pechersky A.V.: Osaline vanusega seotud androgeenidefitsiit. - SPb.: SPbMAPO, 2005
Ed. Yu.A. Eršov; Rec. MITTE. Kuzmenko: Üldine keemia. Biofüüsikaline keemia. Biogeensete elementide keemia. - M.: Kõrgkool, 2005
T.L. Aleinikova ja teised; Ed. E.S. Severina; Arvustaja: D.M. Nikulina, Z.I. Mikašenovitš, L.M. Pustovalova: Biokeemia. - M.: GEOTAR-MED, 2005
Tyukavkina N.A.: Bioorgaaniline keemia. - M.: Bustard, 2005
Zhizhin GV: keemiliste reaktsioonide ja bioloogiliste populatsioonide isereguleeruvad lained. - Peterburi: Nauka, 2004
Ivanov V.P.: Rakumembraanide valgud ja veresoonte düstoonia inimestel. - Kursk: KSMU KMI, 2004
Taimefüsioloogia Instituut im. K.A. Timirjazevi RAS; Rep. toim. V.V. Kuznetsov: Andrei Lvovitš Kursanov: Elu ja töö. - M.: Nauka, 2004
Komov V.P.: Biokeemia. - M.: Bustard, 2004
Muud seotud tööd, mis võivad teile huvi pakkuda.vshm> |
|||
| 21479. | VALGU AINEVAHETUS | 150.03KB | |
| Lämmastikubilansi on kolme tüüpi: lämmastikubilanss positiivne lämmastikubilanss negatiivne lämmastikubilanss Positiivse lämmastikubilansi korral on lämmastiku sissevõtmine ülekaalus selle vabanemise üle. Neeruhaiguse korral on võimalik valepositiivne lämmastikubilanss, mille korral lämmastiku metabolismi lõpp-produktide kehas esineb viivitus. Negatiivse lämmastikubilansi korral on lämmastiku eritumine ülekaalus selle tarbimise üle. See seisund on võimalik selliste haigustega nagu tuberkuloos, reuma, onkoloogilised ... | |||
| 21481. | LIPIIDIDE AINEVAHETUS JA FUNKTSIOONID | 194,66 KB | |
| Rasvad sisaldavad erinevaid alkohole ja rasvhape. Alkohole esindavad glütserool, sfingosiin ja kolesterool.Inimese kudedes on ülekaalus paarisarvulise süsinikuaatomite arvuga pika ahelaga rasvhapped. Eristada küllastunud ja küllastumata rasvhappeid... | |||
| 385. | SÜSIVESIKUTE STRUKTUUR JA AINEVAHETUS | 148,99 KB | |
| Glükoosi ja glükogeeni struktuur ja bioloogiline roll. Heksoosdifosfaadi rada glükoosi lagundamiseks. Süsivesikute avatud ahela ja tsüklilised vormid Joonisel on glükoosi molekul avatud ahela kujul ja tsüklilise struktuuri kujul. Glükoosi tüüpi heksoosides ühineb esimene süsinikuaatom hapnikuga viienda süsinikuaatomi juures, mille tulemusena moodustub kuueliikmeline ring. | |||
| 7735. | KOMMUNIKATSIOON KUI INFOVAHETUS | 35,98 KB | |
| Umbes 70 protsenti teabest edastatakse suhtlusprotsessis mitteverbaalsete suhtluskanalite kaudu ja ainult 30 protsenti verbaalsete kanalite kaudu. Seetõttu ei saa inimese kohta rohkem öelda mitte sõna, vaid pilk, miimika, plastilised poosid, žestid, kehaliigutused, inimestevaheline distants, riietus ja muud mitteverbaalsed suhtlusvahendid. Seega võib mitteverbaalse suhtluse peamisteks ülesanneteks pidada: psühholoogilise kontakti loomist ja hoidmist, suhtlusprotsessi reguleerimist; sõnalisele tekstile uute tähenduslike varjundite lisamine; sõnade õige tõlgendamine;... | |||
| 6645. | Ainevahetus ja energia (ainevahetus) | 39,88 KB | |
| Ainete sisenemine rakku. Suhkrusoolade ja muude lahuste sisalduse tõttu osmootselt toimeaineid rakke iseloomustab teatud osmootse rõhu olemasolu neis. Ainete kontsentratsiooni erinevust rakus ja väljaspool seda nimetatakse kontsentratsioonigradiendiks. | |||
| 21480. | NULEIINHAPPETE AINEVAHETUS JA FUNKTSIOONID | 116,86 KB | |
| Desoksüribonukleiinhape DNA lämmastikualuseid esindab adeniin guaniin tümiin tsütosiin süsivesik – desoksüriboos. DNA mängib olulist rolli geneetilise teabe säilitamisel. Erinevalt RNA-st on DNA-l kaks polünukleotiidahelat. DNA molekulmass on umbes 109 daltonit. | |||
| 386. | RASVDE JA LIPOIDIDE STRUKTUUR JA AINEVAHETUS | 724,43 KB | |
| Lipiidide koostises on leitud arvukalt ja mitmekesiseid struktuurseid komponente: kõrgemad rasvhapped, alkoholid, aldehüüdid, süsivesikud, lämmastikalused, aminohapped, fosforhape jne. Rasvad moodustavad rasvhapped jaotatakse küllastunud ja küllastumata. Rasvhapped Mõned füsioloogiliselt olulised küllastunud rasvhapped C-aatomite arv Triviaalne nimi Süstemaatiline nimetus Keemiline valemühendused... | |||
| 10730. | Rahvusvaheline tehnoloogiavahetus. Rahvusvaheline teenustekaubandus | 56,4 KB | |
| Transporditeenused maailmaturul. Peamine erinevus seisneb selles, et teenustel ei ole tavaliselt materialiseeritud vormi, kuigi paljud teenused omandavad selle, näiteks: magnetkandjatena arvutiprogrammid erinevad paberile trükitud dokumentatsioonid jne. Erinevalt kaupadest toodetakse ja tarbitakse teenuseid peamiselt üheaegselt ning need ei kuulu ladustamisele. olukord, kus teenuse müüja ja ostja ei liigu üle piiri, üle piiri läheb ainult teenus. | |||
| 4835. | Raua metabolism ja raua metabolismi rikkumine. Hemosederoos | 138,5 KB | |
| Raud on oluline mikroelement, mis osaleb hingamises, hematopoeesis, immunobioloogilistes ja redoksreaktsioonides ning on osa enam kui 100 ensüümist. Raud on hemoglobiini ja müohemoglobiini oluline komponent. Täiskasvanu kehas on umbes 4 g rauda, millest üle poole (umbes 2,5 g) moodustab hemoglobiini raud. | |||
Teema tähendus: Vesi ja selles lahustunud ained loovad organismi sisekeskkonna. Vee-soola homöostaasi olulisemad parameetrid on osmootne rõhk, pH ning rakusisese ja rakuvälise vedeliku maht. Nende parameetrite muutused võivad põhjustada vererõhu muutusi, atsidoosi või alkaloosi, dehüdratsiooni ja kudede turset. Peamised hormoonid, mis osalevad vee-soola ainevahetuse peenregulatsioonis ning toimivad neerude distaalsetes tuubulites ja kogumisjuhades: antidiureetiline hormoon, aldosteroon ja natriureetiline faktor; neerude reniin-angiotensiini süsteem. Just neerudes toimub uriini koostise ja mahu lõplik moodustumine, mis tagab sisekeskkonna regulatsiooni ja püsivuse. Neerud eristuvad intensiivse energia metabolismiga, mis on seotud vajadusega uriini moodustumise ajal märkimisväärses koguses ainete aktiivseks transmembraanseks transportimiseks.
Uriini biokeemiline analüüs annab aimu neerude funktsionaalsest seisundist, ainevahetusest erinevates organites ja organismis tervikuna, aitab selgitada patoloogilise protsessi olemust ja annab võimaluse hinnata ravi efektiivsust. .
Tunni eesmärk: uurida vee-soola ainevahetuse parameetrite omadusi ja nende reguleerimise mehhanisme. Ainevahetuse tunnused neerudes. Õppige läbi viima ja hindama biokeemiline analüüs uriin.
Õpilane peab teadma:
1. Uriini moodustumise mehhanism: glomerulaarfiltratsioon, reabsorptsioon ja sekretsioon.
2. Keha veesektsioonide omadused.
3. Keha vedela keskkonna peamised parameetrid.
4. Mis tagab rakusisese vedeliku parameetrite püsivuse?
5. Süsteemid (elundid, ained), mis tagavad rakuvälise vedeliku püsivuse.
6. Tegurid (süsteemid), mis tagavad rakuvälise vedeliku osmootse rõhu ja selle reguleerimise.
7. Tegurid (süsteemid), mis tagavad rakuvälise vedeliku mahu püsivuse ja selle reguleerimise.
8. Tegurid (süsteemid), mis tagavad rakuvälise vedeliku happe-aluselise oleku püsivuse. Neerude roll selles protsessis.
9. Ainevahetuse tunnused neerudes: kõrge metaboolne aktiivsus, Esimene aste kreatiini süntees, intensiivse glükoneogeneesi (isoensüümide) roll, D3-vitamiini aktiveerimine.
10. Üldised omadused uriin (kogus päevas - diurees, tihedus, värvus, läbipaistvus), keemiline koostis uriin. Uriini patoloogilised komponendid.
Õpilane peab suutma:
1. Viige läbi uriini põhikomponentide kvalitatiivne määramine.
2. Hinnake uriini biokeemilist analüüsi.
Õpilane peab saama idee:
Mõnede patoloogiliste seisundite kohta, millega kaasnevad muutused uriini biokeemilistes parameetrites (proteinuuria, hematuuria, glükosuuria, ketonuuria, bilirubinuuria, porfürinuuria) .
Teema uurimiseks vajalik teave põhidistsipliinidest:
1. Neeru struktuur, nefron.
2. Uriini moodustumise mehhanismid.
Enesekoolituse ülesanded:
Tutvuge teema materjaliga vastavalt sihtküsimustele ("õpilane peab teadma") ja täitke kirjalikult järgmised ülesanded:
1. Vaadake histoloogia käiku. Pidage meeles nefroni struktuuri. Pange tähele proksimaalset tuubulit, distaalset keerdtorukest, kogumiskanalit, vaskulaarset glomerulit, jukstaglomerulaarset aparaati.
2. Vaadake normaalse füsioloogia kulgu. Pidage meeles uriini moodustumise mehhanismi: filtreerimine glomerulites, reabsorptsioon tuubulites koos sekundaarse uriini ja sekretsiooni moodustumisega.
3. Rakuvälise vedeliku osmootse rõhu ja mahu reguleerimine on seotud peamiselt naatriumi- ja veeioonide sisalduse reguleerimisega ekstratsellulaarses vedelikus.
Nimetage selle määrusega seotud hormoonid. Kirjeldage nende toimet vastavalt skeemile: hormoonide sekretsiooni põhjus; sihtorgan (rakud); nende toimemehhanism nendes rakkudes; nende tegevuse lõplik mõju.
Pange oma teadmised proovile:
A. Vasopressiin(kõik õiged, välja arvatud üks):
a. sünteesitakse hüpotalamuse neuronites; b. eritub osmootse rõhu tõusuga; sisse. suurendab primaarsest uriinist vee reabsorptsiooni kiirust neerutuubulites; g) suurendab naatriumioonide reabsorptsiooni neerutuubulites; e) vähendab osmootset rõhku e) uriin muutub kontsentreeritumaks.
B. Aldosteroon(kõik õiged, välja arvatud üks):
a. sünteesitakse neerupealiste koores; b. sekreteeritakse, kui naatriumiioonide kontsentratsioon veres väheneb; sisse. neerutuubulites suurendab naatriumioonide reabsorptsiooni; d) uriin muutub kontsentreeritumaks.
e) Peamine sekretsiooni reguleerimise mehhanism on neerude areniin-angiotensiivne süsteem.
B. Natriureetiline tegur(kõik õiged, välja arvatud üks):
a. sünteesitakse aatriumi rakkude alustes; b. sekretsiooni stiimul - vererõhu tõus; sisse. suurendab glomerulite filtreerimisvõimet; d) suurendab uriini moodustumist; e. Uriin muutub vähem kontsentreerituks.
4. Joonistage skeem, mis illustreerib reniin-angiotensiivse süsteemi rolli aldosterooni ja vasopressiini sekretsiooni reguleerimisel.
5. Rakuvälise vedeliku happe-aluse tasakaalu püsivust säilitavad vere puhversüsteemid; muutus kopsuventilatsioonis ja hapete (H +) eritumise kiirus neerude kaudu.
Pidage meeles vere puhversüsteeme (baasvesinikkarbonaat)!
Pange oma teadmised proovile:
Loomset päritolu toit on oma olemuselt happeline (erinevalt toidust peamiselt fosfaatide tõttu taimset päritolu). Kuidas muutub uriini pH inimesel, kes kasutab peamiselt loomset päritolu toitu:
a. lähemal pH 7,0-le; b.pn umbes 5.; sisse. pH umbes 8,0.
6. Vasta küsimustele:
A. Kuidas seletada neerude poolt tarbitava hapniku suurt osakaalu (10%);
B. Glükoneogeneesi kõrge intensiivsus;
B. Neerude roll kaltsiumi metabolismis.
7. Nefronite üks peamisi ülesandeid on õiges koguses kasulike ainete tagasiimendamine verest ja ainevahetuse lõpp-produktide eemaldamine verest.
Tee laud Uriini biokeemilised näitajad:
Auditooriumitöö.
Laboratoorsed tööd:
Viia läbi rida kvalitatiivseid reaktsioone erinevate patsientide uriiniproovides. Tehke biokeemilise analüüsi tulemuste põhjal järeldus metaboolsete protsesside seisundi kohta.
pH määramine.
Töö käik: indikaatorpaberi keskele kantakse 1-2 tilka uriini ning ühe värvilise riba värvi muutmisega, mis langeb kokku kontrollriba värviga, saadakse uuritava uriini pH. kindlaks määratud. Normaalne pH 4,6 - 7,0
2. Kvalitatiivne reaktsioon valkudele. Normaalne uriin ei sisalda valku (normaalsete reaktsioonide käigus ei tuvastata jälgi). Mõne patoloogilise seisundi korral võib uriinis esineda valku - proteinuuria.
Edusammud: 1-2 ml uriinile lisada 3-4 tilka värskelt valmistatud 20% sulfasalitsüülhappe lahust. Valgu juuresolekul ilmub valge sade või hägusus.
3. Kvalitatiivne reaktsioon glükoosile (Fehlingi reaktsioon).
Töö käik: Lisage 10 tilka uriinile 10 tilka Fehlingi reaktiivi. Kuumuta keemiseni. Glükoosi juuresolekul ilmub punane värv. Võrrelge tulemusi normiga. Tavaliselt ei tuvastata kvalitatiivsete reaktsioonide abil glükoosi jälgi uriinis. Tavaliselt ei ole uriinis glükoosi. Mõne patoloogilise seisundi korral ilmneb glükoos uriinis. glükosuuria.
Määramise saab läbi viia testriba (indikaatorpaber) abil /
Ketoonkehade tuvastamine
Töö käik: kandke slaidile tilk uriini, tilk 10% naatriumhüdroksiidi lahust ja tilk värskelt valmistatud 10% naatriumnitroprussiidi lahust. Ilmub punane värv. Valage 3 tilka kontsentreeritud äädikhapet - ilmub kirsivärv.
Tavaliselt ketokehad uriinis puuduvad. Mõne patoloogilise seisundi korral ilmuvad uriinis ketokehad - ketonuuria.
Lahendage probleeme ise, vastake küsimustele:
1. Ekstratsellulaarse vedeliku osmootne rõhk on tõusnud. Kirjeldage skemaatilisel kujul sündmuste jada, mis viivad selle vähenemiseni.
2. Kuidas muutub aldosterooni tootmine, kui vasopressiini liigne tootmine toob kaasa osmootse rõhu olulise languse.
3. Kirjeldage sündmuste jada (diagrammi kujul), mille eesmärk on taastada homöostaas koos naatriumkloriidi kontsentratsiooni vähenemisega kudedes.
4. Patsiendil on suhkurtõbi, millega kaasneb ketoneemia. Kuidas reageerib happe-aluse tasakaalu muutustele peamine verepuhvri süsteem – vesinikkarbonaat? Milline on neerude roll KOS-i taastumisel? Kas selle patsiendi uriini pH muutub.
5. Võistlusteks valmistuv sportlane läbib intensiivse treeningu. Kuidas muuta glükoneogeneesi kiirust neerudes (vaidle vastust)? Kas sportlasel on võimalik uriini pH-d muuta; põhjendage vastust)?
6. Patsiendil on luukoes ainevahetushäire tunnused, mis mõjutavad ka hammaste seisundit. Kaltsitoniini ja paratüreoidhormooni tase on füsioloogilise normi piires. Patsient saab D-vitamiini (kolekaltsiferooli) vajalikes kogustes. Tee oletus umbes võimalik põhjus ainevahetushäired.
7. Kaaluge standardvormi " Üldine analüüs uriin” (Tjumeni Riikliku Meditsiiniakadeemia mitmeprofiililine kliinik) ning oskama selgitada biokeemilistes laborites määratud uriini biokeemiliste komponentide füsioloogilist rolli ja diagnostilist tähtsust. Pidage meeles, et uriini biokeemilised parameetrid on normaalsed.
Vee ainevahetuse reguleerimine toimub neurohumoraalsel teel, eelkõige kesknärvisüsteemi erinevate osade poolt: ajukoor, vahe- ja piklikaju, sümpaatilised ja parasümpaatilised ganglionid. Kaasatud on ka paljud endokriinsed näärmed. Hormoonide toime seisneb sel juhul selles, et nad muudavad rakumembraanide läbilaskvust vee jaoks, tagades selle vabanemise ehk tagasisorptsiooni.Keha veevajadust reguleerib janu. Juba esimeste vere paksenemise tunnuste ilmnemisel tekib ajukoore teatud osade reflektoorse ergastuse tagajärjel janu. Tarbitav vesi imendub sel juhul läbi sooleseina ning selle liig ei põhjusta vere vedeldamist. . Alates verd, liigub see kiiresti lahtise sidekoe, maksa, naha jne rakkudevahelistesse ruumidesse.Need koed toimivad kehas vee depoona.Üksikutel katioonidel on teatav mõju vee sissevõtule ja kudedest vabanemisele. Na + ioonid aitavad kaasa valkude sidumisele kolloidosakeste kaudu, K + ja Ca 2+ ioonid stimuleerivad vee väljutamist kehast.
Seega soodustab neurohüpofüüsi vasopressiin (antidiureetiline hormoon) vee tagasisorptsiooni primaarsest uriinist, vähendades viimase eritumist organismist. Neerupealise koore hormoonid - aldosteroon, deoksükortikosterool - aitavad kaasa naatriumi säilimisele organismis ja kuna naatriumkatioonid suurendavad kudede hüdratatsiooni, siis säilib neis ka vesi. Teised hormoonid stimuleerivad vee eritumist neerude kaudu: türoksiin – hormoon kilpnääre, parathormoon – hormoon kõrvalkilpnääre, androgeenid ja östrogeenid – sugunäärmete hormoonid Kilpnäärmehormoonid stimuleerivad vee eritumist läbi higinäärmete Vee hulk kudedes, peamiselt vabas, suureneb neeruhaiguste, südame-veresoonkonna talitlushäirete, valgunälgimise korral. maksafunktsiooni kahjustus (tsirroos). Veesisalduse suurenemine rakkudevahelistes ruumides põhjustab turset. Vasopressiini ebapiisav moodustumine põhjustab diureesi suurenemist, diabeedi insipidus'e haigust. Keha dehüdratsiooni täheldatakse ka aldosterooni ebapiisava moodustumise korral neerupealiste koores.
Vesi ja selles lahustunud ained, sh mineraalsoolad, loovad organismi sisekeskkonna, mille omadused püsivad konstantsena või muutuvad korrapäraselt elundite ja rakkude funktsionaalse seisundi muutumisel.Keha vedela keskkonna peamised parameetrid on osmootne rõhk,pH ja maht.
Ekstratsellulaarse vedeliku osmootne rõhk sõltub suuresti soolast (NaCl), mis selles vedelikus sisaldub suurimas kontsentratsioonis. Seetõttu on osmootse rõhu reguleerimise põhimehhanism seotud kas vee või NaCl vabanemise kiiruse muutumisega, mille tulemusena muutub NaCl kontsentratsioon koevedelikes, mis tähendab, et muutub ka osmootne rõhk. Mahu reguleerimine toimub nii vee kui ka NaCl vabanemise kiiruse samaaegse muutmise teel. Lisaks reguleerib janumehhanism veetarbimist. pH reguleerimise tagab hapete või leeliste selektiivne eritumine uriiniga; Uriini pH võib sellest olenevalt varieeruda vahemikus 4,6 kuni 8,0. Patoloogilised seisundid, nagu kudede dehüdratsioon või tursed, vererõhu tõus või langus, šokk, atsidoos ja alkaloos, on seotud vee-soola homöostaasi rikkumisega.
Osmootse rõhu ja rakuvälise vedeliku mahu reguleerimine. Vee ja NaCl eritumist neerude kaudu reguleerivad antidiureetiline hormoon ja aldosteroon.
Antidiureetiline hormoon (vasopressiin). Vasopressiin sünteesitakse hüpotalamuse neuronites. Hüpotalamuse osmoretseptorid stimuleerivad vasopressiini vabanemist sekretoorsetest graanulitest koos koevedeliku osmootse rõhu tõusuga. Vasopressiin suurendab primaarsest uriinist vee reabsorptsiooni kiirust ja vähendab seeläbi diureesi. Uriin muutub kontsentreeritumaks. Nii hoiab antidiureetiline hormoon kehas vajaliku vedelikumahu, mõjutamata vabaneva NaCl kogust. Väheneb rakuvälise vedeliku osmootne rõhk, s.t elimineeritakse vasopressiini vabanemist põhjustanud stiimul.Mõnede hüpotalamust või hüpofüüsi kahjustavate haiguste korral (kasvajad, vigastused, infektsioonid) vasopressiini süntees ja sekretsioon väheneb ja areneb. diabeet insipidus.
Lisaks diureesi vähendamisele põhjustab vasopressiin ka arterioolide ja kapillaaride ahenemist (sellest ka nimi) ja sellest tulenevalt vererõhu tõusu.
Aldosteroon. Seda steroidhormooni toodetakse neerupealiste koores. Sekretsioon suureneb koos NaCl kontsentratsiooni vähenemisega veres. Neerudes suurendab aldosteroon Na + (ja koos sellega C1) reabsorptsiooni kiirust nefronituubulites, mis põhjustab NaCl peetust organismis. See kõrvaldab stiimuli, mis põhjustas aldosterooni sekretsiooni.Aldosterooni liigne sekretsioon põhjustab vastavalt NaCl liigset retentsiooni ja rakuvälise vedeliku osmootse rõhu tõusu. Ja see toimib signaalina vasopressiini vabanemiseks, mis kiirendab vee reabsorptsiooni neerudes. Selle tulemusena koguneb kehasse nii NaCl kui vesi; rakuvälise vedeliku maht suureneb, säilitades samal ajal normaalse osmootse rõhu.
Reniini-angiotensiini süsteem. See süsteem toimib aldosterooni sekretsiooni reguleerimise peamise mehhanismina; sellest sõltub ka vasopressiini sekretsioon Reniin on proteolüütiline ensüüm, mis sünteesitakse neeruglomeruluse aferentset arteriooli ümbritsevates jukstaglomerulaarrakkudes.
Reniini-angiotensiini süsteemil on oluline roll veremahu taastamisel, mis võib väheneda verejooksu, tugeva oksendamise, kõhulahtisuse (diarröa) ja higistamise tagajärjel. Oma osa mängib angiotensiin II toimel tekkinud vasokonstriktsioon erakorraline meede vererõhu säilitamiseks. Siis jäävad organismis tavapärasest suuremal määral joogi ja toiduga kaasnev vesi ja NaCl, mis tagab veremahu ja rõhu taastumise. Pärast seda lakkab reniini vabanemine, veres juba esinevad regulatoorsed ained hävivad ja süsteem naaseb algsesse olekusse.
Tsirkuleeriva vedeliku mahu märkimisväärne vähenemine võib põhjustada kudede verevarustuse ohtlikku rikkumist, enne kui regulatsioonisüsteemid taastavad rõhu ja veremahu. Samal ajal on häiritud kõigi elundite ja ennekõike aju funktsioonid; tekib seisund, mida nimetatakse šokiks. Šoki (nagu ka tursete) tekkes on oluline roll vedeliku ja albumiini normaalse jaotumise muutumisel vereringe ja rakkudevahelise ruumi vahel.Vesopressiin ja aldosteroon osalevad vee-soola tasakaalu reguleerimises, toimivad nefronituubulite tasemel - need muudavad primaarsete uriinikomponentide reabsorptsiooni kiirust.
Vee-soola ainevahetus ja seedemahlade eritumine. Kõigi seedenäärmete igapäevase sekretsiooni maht on üsna suur. Normaalsetes tingimustes imendub nende vedelike vesi soolestikus uuesti; tugev oksendamine ja kõhulahtisus võivad põhjustada rakuvälise vedeliku mahu olulist vähenemist ja kudede dehüdratsiooni. Märkimisväärne vedelikukaotus seedemahladega toob kaasa albumiini kontsentratsiooni suurenemise vereplasmas ja rakkudevahelises vedelikus, kuna albumiin ei eritu koos saladustega; sel põhjusel suureneb rakkudevahelise vedeliku osmootne rõhk, vesi hakkab rakkudest liikuma rakkudevahelisse vedelikku ja raku funktsioonid on häiritud. Ka ekstratsellulaarse vedeliku kõrge osmootne rõhk viib uriini tootmise vähenemiseni või isegi lakkamiseni. , ja kui vett ja soolasid väljastpoolt ei anta, tekib loomal kooma.
Homöostaasi ühe külje säilitamine - keha vee-elektrolüütide tasakaal viiakse läbi neuroendokriinse regulatsiooni abil. Kõrgeim vegetatiivne janukeskus asub ventromediaalses hüpotalamuses. Vee ja elektrolüütide vabanemise reguleerimine toimub peamiselt neerufunktsiooni neurohumoraalse kontrolli kaudu. Erilist rolli selles süsteemis mängivad kaks tihedalt seotud neurohormonaalset mehhanismi – aldosterooni ja (ADH) sekretsiooni. Aldosterooni reguleeriva toime põhisuund on selle inhibeeriv toime kõikidele naatriumi eritumise teedele ja eelkõige neerutuubulitele (natriureemivastane toime). ADH säilitab vedeliku tasakaalu, pärssides otseselt vee eritumist neerude kaudu (antidiureetiline toime). Aldosterooni aktiivsuse ja antidiureetilise toimemehhanismi vahel on pidev ja lähedane seos. Vedelikukaotus stimuleerib aldosterooni sekretsiooni volomoretseptorite kaudu, mille tulemuseks on naatriumi peetus ja ADH kontsentratsiooni tõus. Mõlema süsteemi efektororganid on neerud.
Vee ja naatriumi kadu määra määravad vee-soola metabolismi humoraalse reguleerimise mehhanismid: hüpofüüsi antidiureetiline hormoon, vasopressiin ja neerupealiste hormoon aldosteroon, mis toimivad kõige olulisemale organile, kinnitades vee-soola tasakaalu püsivust. kehas, mis on neerud. ADH toodetakse hüpotalamuse supraoptilistes ja paraventrikulaarsetes tuumades. Hüpofüüsi portaalsüsteemi kaudu siseneb see peptiid hüpofüüsi tagumisse ossa, kontsentreerub seal ja vabaneb verre hüpofüüsi sisenevate närviimpulsside mõjul. ADH sihtmärk on neerude distaalsete tuubulite sein, kus see suurendab depolümeriseeriva hüaluronidaasi tootmist. hüaluroonhape suurendades seeläbi anuma seinte läbilaskvust. Selle tulemusena difundeerub vesi primaarsest uriinist passiivselt neerurakkudesse osmootse gradiendi tõttu keha hüperosmootse rakkudevahelise vedeliku ja hüpoosmolaarse uriini vahel. Neerud läbivad päevas oma veresoonte kaudu umbes 1000 liitrit verd. 180 liitrit primaarset uriini filtreeritakse läbi neerude glomerulite, kuid ainult 1% neerude kaudu filtreeritud vedelikust muutub uriiniks, 6/7 primaarse uriini moodustavast vedelikust läbib kohustusliku reabsorptsiooni koos teiste lahustunud ainetega. see proksimaalsetes tuubulites. Ülejäänud primaarne uriinivesi reabsorbeerub distaalsetes tuubulites. Neis toimub primaarse uriini moodustumine mahu ja koostise osas.
Rakuvälises vedelikus reguleerivad osmootset rõhku neerud, mis võivad eritada uriini naatriumkloriidi kontsentratsiooniga, mis jääb vahemikku 340 mmol/l. Naatriumkloriidi vaese uriini eraldumisel suureneb soolapeetuse tõttu osmootne rõhk ja soola kiire vabanemisega langeb.
Uriini kontsentratsiooni kontrollivad hormoonid: vasopressiin (antidiureetiline hormoon), suurendades vee vastupidist imendumist, suurendab soola kontsentratsiooni uriinis, aldosteroon stimuleerib naatriumi vastupidist imendumist. Nende hormoonide tootmine ja sekretsioon sõltub osmootsest rõhust ja naatriumi kontsentratsioonist rakuvälises vedelikus. Plasma soolade kontsentratsiooni vähenemisega suureneb aldosterooni tootmine ja naatriumi peetus, suurenedes vasopressiini tootmine suureneb ja aldosterooni tootmine väheneb. See suurendab vee tagasiimendumist ja naatriumi kadu ning aitab vähendada osmootset rõhku. Lisaks põhjustab osmootse rõhu tõus janu, mis suurendab veetarbimist. Vasopressiini moodustumise ja janutunde signaalid käivitavad hüpotalamuses osmoretseptorid.
Rakkude mahu reguleerimine ja ioonide kontsentratsioon rakkudes on energiast sõltuvad protsessid, sealhulgas naatriumi ja kaaliumi aktiivne transport läbi rakumembraanide. Aktiivsete transpordisüsteemide energiaallikaks, nagu peaaegu iga raku energiakulu puhul, on ATP vahetus. Juhtiv ensüüm, naatrium-kaalium-ATPaas, annab rakkudele võime pumbata naatriumi ja kaaliumi. See ensüüm nõuab magneesiumi ja lisaks on maksimaalse aktiivsuse saavutamiseks vajalik nii naatriumi kui ka kaaliumi samaaegne olemasolu. Kaaliumi ja teiste ioonide erinevate kontsentratsioonide olemasolu rakumembraani vastaskülgedel on üks tagajärgi, et membraani ulatuses tekivad elektripotentsiaalide erinevused.
Naatriumpumba töö tagamiseks kulub kuni 1/3 kogu skeletilihasrakkude salvestatud energiast. Hüpoksia või mis tahes metabolismi inhibiitorite sekkumise korral rakk paisub. Turse mehhanism on naatriumi- ja kloriidioonide sisenemine rakku; see toob kaasa intratsellulaarse osmolaarsuse suurenemise, mis omakorda suurendab veesisaldust, kui see järgneb lahustunud ainele. Samaaegne kaaliumikadu ei ole samaväärne naatriumi tarbimisega ja seetõttu on tulemuseks veesisalduse suurenemine.
Ekstratsellulaarse vedeliku efektiivne osmootne kontsentratsioon (toonisus, osmolaarsus) muutub peaaegu paralleelselt selles sisalduva naatriumi kontsentratsiooniga, mis koos anioonidega annab vähemalt 90% selle osmootsest aktiivsusest. Kaaliumi ja kaltsiumi kõikumised (isegi patoloogilistes tingimustes) ei ületa paari milliekvivalenti 1 liitri kohta ega mõjuta oluliselt osmootset rõhku.
Ekstratsellulaarse vedeliku hüpoelektrolüteemia (hüpoosmia, hüpoosmolaarsus, hüpotoonilisus) on osmootse kontsentratsiooni langus alla 300 mosm / l. See vastab naatriumi kontsentratsiooni langusele alla 135 mmol/L. Hüperelektrolüteemia (hüperosmolaarsus, hüpertoonilisus) on osmootse kontsentratsiooni ületamine 330 mosm / l ja naatriumi kontsentratsioon 155 mmol / l.
Suured vedelikumahtude kõikumised keha sektorites on tingitud keerulistest bioloogilistest protsessidest, mis järgivad füüsikalisi ja keemilisi seadusi. Sel juhul on suur tähtsus elektrilise neutraalsuse printsiibil, mis seisneb selles, et positiivsete laengute summa kõigis veeruumides on võrdne negatiivsete laengute summaga. Pidevalt toimuvate muutustega elektrolüütide kontsentratsioonis vesikeskkonnas kaasneb elektriliste potentsiaalide muutus koos järgneva taastumisega. Dünaamilises tasakaalus moodustuvad bioloogiliste membraanide mõlemal küljel stabiilsed katioonide ja anioonide kontsentratsioonid. Siiski tuleb märkida, et elektrolüüdid ei ole ainsad osmootselt aktiivsed komponendid keha vedelas keskkonnas, mis toiduga kaasas on. Süsivesikute ja rasvade oksüdeerumine viib tavaliselt süsihappegaasi ja vee moodustumiseni, mis võivad kopsude kaudu lihtsalt väljutada. Aminohapete oksüdeerumisel moodustub ammoniaak ja uurea. Ammoniaagi muundumine karbamiidiks annab inimkehale ühe võõrutusmehhanismidest, kuid samal ajal muutuvad lenduvad ühendid, mis võivad kopsude kaudu eemaldada, mittelenduvateks, mis peaksid juba neerude kaudu erituma.
Vee ja elektrolüütide, toitainete, hapniku ja süsihappegaasi ning muude ainevahetuse lõpp-produktide vahetus on peamiselt tingitud difusioonist. Kapillaarvesi vahetab vett interstitsiaalse koega mitu korda sekundis. Lipiidide lahustuvuse tõttu difundeeruvad hapnik ja süsinikdioksiid vabalt läbi kõigi kapillaarmembraanide; samal ajal arvatakse, et vesi ja elektrolüüdid läbivad endoteelimembraani väikseimaid poore.
7. Vee ainevahetuse häirete klassifitseerimise põhimõtted ja peamised liigid.
Tuleb märkida, et vee- ja elektrolüütide tasakaaluhäiretel puudub ühtne üldtunnustatud klassifikatsioon. Igat tüüpi häired, sõltuvalt vee mahu muutusest, jagunevad tavaliselt: rakuvälise vedeliku mahu suurenemisega - veebilanss on positiivne (hüperhüdratsioon ja tursed); rakuvälise vedeliku mahu vähenemisega - negatiivne veebilanss (dehüdratsioon). Hamburger et al. (1952) tegi ettepaneku jagada kõik need vormid rakuvälisteks ja rakkudevaheliseks. Vee koguhulga üle- ja vähenemist arvestatakse alati seoses naatriumi kontsentratsiooniga rakuvälises vedelikus (selle osmolaarsusega). Sõltuvalt osmootse kontsentratsiooni muutusest jagatakse hüper- ja dehüdratsioon kolme tüüpi: isoosmolaarne, hüpoosmolaarne ja hüperosmolaarne.
Liigne vee kogunemine kehas (hüperhüdratsioon, hüperhüdria).
Isotooniline hüperhüdratsioon tähistab ekstratsellulaarse vedeliku mahu suurenemist ilma osmootset rõhku häirimata. Sel juhul ei toimu vedeliku ümberjaotumist intra- ja ekstratsellulaarsete sektorite vahel. Vee kogumahu suurenemine organismis on tingitud rakuvälisest vedelikust. Selline seisund võib olla südamepuudulikkuse, nefrootilise sündroomi hüpoproteineemia tagajärg, kui tsirkuleeriva vere maht jääb vedela osa liikumise tõttu interstitsiaalsesse segmenti konstantseks (ilmub palpeeritav jäsemete turse, võib tekkida kopsuturse). Viimane võib olla seotud tõsiste tüsistustega parenteraalne manustamine terapeutilistel eesmärkidel kasutatavad vedelikud, suure koguse soolalahuse või Ringeri lahuse infusioon katses või patsientidele operatsioonijärgsel perioodil.
Hüpoosmolaarne ülehüdratsioon või veemürgitus vee liigsest kogunemisest ilma vastava elektrolüütide peetuseta, vedeliku eritumise häire tõttu neerupuudulikkus või antidiureetilise hormooni ebapiisav sekretsioon. Katses saab seda rikkumist reprodutseerida hüpoosmootse lahuse peritoneaaldialüüsiga. Loomadel tekib veemürgitus kergesti ka pärast ADH sisseviimist või neerupealiste eemaldamist veega koormamisel. Tervetel loomadel tekkis veemürgitus 4-6 tundi pärast vee allaneelamist annuses 50 ml/kg iga 30 minuti järel. Tekivad oksendamine, treemor, kloonilised ja toonilised krambid. Elektrolüütide, valkude ja hemoglobiini kontsentratsioon veres väheneb järsult, plasma maht suureneb, vere reaktsioon ei muutu. Jätkuv infusioon võib põhjustada kooma ja loomade surma.
Veemürgituse korral väheneb ekstratsellulaarse vedeliku osmootne kontsentratsioon selle lahjendamise tõttu liigse veega, tekib hüponatreemia. Osmootne gradient "interstitsiumi" ja rakkude vahel põhjustab osa rakkudevahelise vee liikumise rakkudesse ja nende turse. Rakuvee maht võib suureneda 15%.
AT kliiniline praktika koos veemürgistuse nähtustega esinevad juhtudel, kui vee tarbimine ületab neerude võimet seda eritada. Pärast patsiendile 5 või enama liitri vee joomist päevas tekivad vasikatel peavalud, apaatia, iiveldus ja krambid. Veemürgitus võib tekkida liigse veetarbimisega, kui suureneb ADH ja oliguuria tootmine. Pärast vigastusi, suurte kirurgiliste operatsioonide ajal, verekaotus, anesteetikumide, eriti morfiini sisseviimine, kestab oliguuria tavaliselt vähemalt 1-2 päeva. Veemürgitus võib tekkida suure koguse isotoonilise glükoosilahuse intravenoossel infusioonil, mis rakkudes kiiresti ära kulub, ja süstitava vedeliku kontsentratsioon langeb. Samuti on ohtlik sisse viia suurtes kogustes vett piiratud neerufunktsiooniga, mis juhtub šokiga, neeruhaigus anuuria ja oliguuriaga, diabeedi insipiduse ravi ADH ravimitega. Veemürgistuse oht tekib imikute kõhulahtisuse tõttu toksikoosi ravi ajal liigsest sooladeta vee sissetoomisest. Sageli korduvate klistiiride korral esineb mõnikord liigset kastmist.
Terapeutiline toime hüpoosmolaarse hüperhüdria tingimustes peaks olema suunatud liigse vee eemaldamisele ja rakuvälise vedeliku osmootse kontsentratsiooni taastamisele. Kui liig oli seotud anuuriasümptomitega patsiendile liiga suure vee manustamisega, annab tehisneeru kasutamine kiire ravitoime. Taastumine normaalne tase osmootne rõhk soola sisseviimisega on lubatud ainult siis, kui soola koguhulk kehas on vähenenud ja ilmnevad veemürgituse tunnused.
Hüperosomaalne ülehüdratsioon mis väljendub vedeliku mahu suurenemises rakuvälises ruumis koos hüpernatreemiast tingitud osmootse rõhu samaaegse suurenemisega. Häirete tekkemehhanism on järgmine: naatriumipeetusega ei kaasne piisavas mahus veepeetust, rakuväline vedelik osutub hüpertoonseks ja vesi liigub rakkudest rakuvälistesse ruumidesse kuni osmootse tasakaaluni. Rikkumise põhjused on mitmekesised: Cushingi või Kohni sündroom, merevee joomine, traumaatiline ajukahjustus. Kui hüperosmolaarse hüperhüdratsiooni seisund püsib pikka aega, võib tekkida kesknärvisüsteemi rakusurm.
Rakkude dehüdratsioon katsetingimustes toimub hüpertooniliste elektrolüütide lahuste sisseviimisel kogustes, mis ületavad neerude kaudu piisavalt kiire eritumise võimalust. Inimestel tekib sarnane häire, kui nad on sunnitud jooma merevett. Toimub vee liikumine rakkudest rakuvälisesse ruumi, mida tuntakse tugeva janutundena. Mõnel juhul kaasneb turse tekkega hüperosmolaarne hüperhüdria.
Vee kogumahu vähenemine (dehüdratsioon, hüpohüdria, dehüdratsioon, ekssikoos) esineb ka rakuvälise vedeliku osmootse kontsentratsiooni vähenemise või suurenemisega. Dehüdratsiooni oht on verehüüvete oht. Tõsised sümptomid dehüdratsioon tekib pärast umbes ühe kolmandiku rakuvälise vee kaotust.
Hüpoosmolaarne dehüdratsioon areneb nendel juhtudel, kui keha kaotab palju elektrolüüte sisaldavat vedelikku ja kaotus kompenseeritakse väiksema veehulgaga ilma soola lisamata. See seisund esineb korduva oksendamise, kõhulahtisuse, suurenenud higistamise, hüpoaldosteronismi, polüuuria (diabeetsus ja suhkurtõbi) korral, kui veekaotust (hüpotoonilised lahused) täiendatakse osaliselt ilma soolata joomisega. Hüpoosmootilisest ekstratsellulaarsest ruumist tormab osa vedelikust rakkudesse. Seega kaasneb ekssikoosiga, mis areneb soolapuuduse tagajärjel, rakusisene turse. Janu tunnet pole. Veekaotusega veres kaasneb hematokriti tõus, hemoglobiini ja valkude kontsentratsiooni tõus. Vere ammendumine veega ning sellega kaasnev plasmamahu vähenemine ja viskoossuse suurenemine häirib oluliselt vereringet ning põhjustab mõnikord kollapsi ja surma. Minutimahu vähenemine põhjustab ka neerupuudulikkust. Filtreerimismaht langeb järsult ja tekib oliguuria. Uriinis praktiliselt puudub naatriumkloriid, mida soodustab aldosterooni suurenenud sekretsioon, mis on tingitud hulgiretseptorite ergastusest. Jääklämmastiku sisaldus veres suureneb. Võib esineda dehüdratsiooni väliseid märke – turgori vähenemine ja naha kortsumine. Sageli on peavalu, isutus. Dehüdratsiooniga lastel ilmnevad kiiresti apaatia, letargia ja lihasnõrkus.
Hüpoosmolaarse hüdratatsiooni ajal tekkinud vee ja elektrolüütide defitsiit on soovitatav asendada erinevaid elektrolüüte sisaldava isoosmootse või hüpoosmootse vedeliku sisseviimisega. Kui piisav suukaudne vee tarbimine ei ole võimalik, tuleb paratamatu veekaotus läbi naha, kopsude ja neerude kompenseerida 0,9% naatriumkloriidi lahuse intravenoosse infusiooniga. Juba tekkinud puuduse korral suurendatakse süstitavat mahtu, mitte üle 3 liitri päevas. Hüpertoonilist soolalahust tohib manustada ainult erandjuhtudel, kui esineb vere elektrolüütide kontsentratsiooni languse kahjulik mõju, kui neerud ei säilita naatriumi ja palju läheb muul viisil kaduma, vastasel juhul võib liigse naatriumi manustamine suurendada dehüdratsiooni. . Hüperkloreemilise atsidoosi vältimiseks koos neerude eritusfunktsiooni vähenemisega on mõistlik lisada naatriumkloriidi asemel piimhappesool.
Hüperosmolaarne dehüdratsioon areneb selle tarbimist ületava veekao ja endogeense moodustumise tagajärjel ilma naatriumi kadumiseta. Veekadu sellisel kujul toimub vähese elektrolüütide kaoga. See võib ilmneda suurenenud higistamise, hüperventilatsiooni, kõhulahtisuse, polüuuria korral, kui kaotatud vedelikku ei kompenseerita joomisega. Suur veekadu uriinis tekib nn osmootse (või lahjendava) diureesi korral, kui neerude kaudu eritub palju glükoosi, uureat või muid lämmastikku sisaldavaid aineid, mis suurendavad primaarse uriini kontsentratsiooni ja takistavad vee tagasiimendumist. Veekadu ületab sellistel juhtudel naatriumi kadu. Piiratud vee manustamine neelamishäiretega patsientidele, samuti janu mahasurumiseks ajuhaiguste korral, koomas, eakatel, enneaegsetel vastsündinutel, ajukahjustusega imikutel jne. Esimese elupäeva vastsündinud mõnikord esineb hüperosmolaarne eksikoos vähese piimatarbimise tõttu ("janupalavik"). Hüperosmolaarne dehüdratsioon esineb imikutel palju kergemini kui täiskasvanutel. Imikueas võib palaviku, kerge atsidoosi ja muude hüperventilatsiooni juhtude korral kopsude kaudu kaduda suures koguses vett, peaaegu ilma elektrolüütideta. Imikutel võib vee ja elektrolüütide tasakaalu rikkumine tekkida ka neerude vähearenenud keskendumisvõime tõttu. Elektrolüütide peetus tekib lapse kehas palju kergemini, eriti hüpertoonilise või isotoonilise lahuse üledoosi korral. Väikelastel on minimaalne kohustuslik vee eritumine (läbi neerude, kopsude ja naha) pindalaühiku kohta ligikaudu kaks korda suurem kui täiskasvanutel.
Veekao ülekaal elektrolüütide vabanemise üle toob kaasa rakuvälise vedeliku osmootse kontsentratsiooni suurenemise ja vee liikumise rakkudest rakuvälisesse ruumi. Seega aeglustub vere hüübimine. Rakuvälise ruumi mahu vähenemine stimuleerib aldosterooni sekretsiooni. See säilitab sisekeskkonna hüperosmolaarsuse ja vedeliku mahu taastumise tänu suurenenud ADH tootmisele, mis piirab vee kadu neerude kaudu. Ekstratsellulaarse vedeliku hüperosmolaarsus vähendab ka vee eritumist neeruväliste kanalite kaudu. Hüperosmolaarsuse kahjulikku mõju seostatakse rakkude dehüdratsiooniga, mis põhjustab piinavat janutunnet, valkude suurenenud lagunemist ja palavikku. Närvirakkude kadumine toob kaasa psüühikahäired (teadvuse hägusus), hingamishäired. Hüperosmolaarset tüüpi dehüdratsiooniga kaasneb ka kehakaalu langus, naha ja limaskestade kuivus, oliguuria, vere hüübimise tunnused ja vere osmootse kontsentratsiooni tõus. Janu mehhanismi pärssimine ja mõõduka ekstratsellulaarse hüperosmolaarsuse kujunemine katses saavutati süstimisega kassidel hüpotalamuse suprooptilistesse tuumadesse ja rottidel ventromediaalsetesse tuumadesse. Inimese kehavedeliku veepuuduse ja isotoonilisuse taastamine saavutatakse peamiselt aluselisi elektrolüüte sisaldava hüpotoonilise glükoosilahuse sisseviimisega.
Isotooniline dehüdratsioon võib tekkida naatriumi ebanormaalselt suurenenud eritumisel, kõige sagedamini koos näärmete sekretsiooniga seedetrakti(isoosmolaarsed eritised, mille päevane maht on kuni 65% kogu rakuvälise vedeliku mahust). Nende isotooniliste vedelike kadu ei too kaasa rakusisese mahu muutust (kõik kaod tulenevad rakuvälisest mahust). Nende põhjused on korduv oksendamine, kõhulahtisus, kaotus fistuli kaudu, suurte transudaatide moodustumine (astsiit, pleuraefusioon), vere- ja plasmakadu põletuste ajal, peritoniit, pankreatiit.