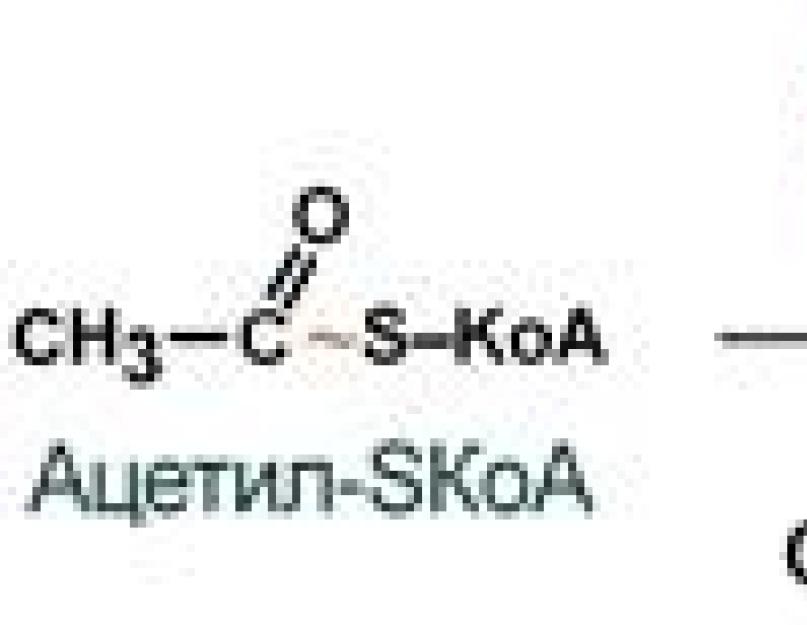
Rasvade biosüntees
Sisaldab rasvhapete ja triatsüülglütseriidide (tegelikult rasvade) biosünteesi.
Rasvhapete biosüntees toimub kõrge glükoosikontsentratsiooni korral veres, peamiselt maksas ja rasvkoes. Sel perioodil aktiveeritakse glükolüüs, mille tulemusena moodustuvad rasvhapete sünteesiks vajalikud substraadid: atsetüül-CoA, ATP, (NADP H + H +) ja teised. Rasvhapete biosünteesi peamine ehitusplokk on atsetüül-CoA ja peamine lõpptoode on palmitiinhape C15H31COOH.
Teised rasvhapped tekivad reeglina palmitiinhappe molekuli modifitseerimisel – ahela pikendamisel ja dehüdrogeenimisel. Viimasel juhul tekivad küllastumata happed.
Palmitiinhappe süntees ei toimu mitokondrites, kus toimub rasvhapete katabolism, vaid tsütosoolis. Selle biosünteesi peamine ensüüm on multiensüümide kompleks palmmetüülsüntetaas. Kuna mitokondri membraan on atsetüül-CoA-le mitteläbilaskev, esialgne etapp biosüntees on atsetüül-CoA ülekandmine läbi mitokondriaalse membraani tsitraat-püruvaadi süstikumehhanismi kaudu.
On teada, et Krebsi tsükli esimene reaktsioon on atsetüül-CoA kondenseerumine oksaloäädikhappega (oksaloatsetaat), moodustades tsitraadi (sidrunhape). Osa moodustunud tsitraadiioonidest ei osale Krebsi tsükli edasistes reaktsioonides, vaid kandub läbi mitokondriaalse membraani tsütosooli, kus tsitraatlüaasi juuresolekul ning ATP ja HS-KoA osalusel see uuesti moodustub. atsetüül-CoA ja PAA:
Tsitraat + HS-KoA + ATP → oksaloatsetaat + atsetüül-CoA + ADP + H 3 RO 4
Oksaloatsetaadi tagastamine mitokondritesse toimub kahe vahendaja - malaadi ja püruvaadi - abil.
Oksaloatsetaadi redutseerimine malaadiks tsütosoolis on osa malaat-aspartaadi süstikumehhanismist redutseeritud (NAD ∙ H + H +) ülekandmiseks tsütosoolist mitokondritesse:
Oksaloatsetaat + NAD∙H + H + ↔ Malaat + NAD
Saadud malaati aga ei transpordita membraanist/läbi membraani, vaid see oksüdeeritakse kohe koos samaaegse dekarboksüülimisega püruvaadiks:
Malaat + NADP + → püruvaat + CO 2 + NADP∙H + H +
Kõik kirjeldatud teisendused on näidatud diagrammil:
Seega kaasneb ühe atsetüül-CoA molekuli ülekandmisega mitokondritest tsütosooli ühe redutseeritud vormi (NADP H + H +) molekuli moodustumine, mis on vajalik paljude biosünteesi jaoks, ja püruvaadi moodustumine, mis hajub mitokondritesse, karboksüülitakse seejärel oksaloatsetaadiks.
Palmitiinhappe tegelik süntees algab atsetüül-CoA karboksüülimisega. See reaktsioon toimub ensüümi juuresolekul, mille proteesrühmaks on biotiin:
See reaktsioon on rasvhapete sünteesis võtmetähtsusega. Edasised transformatsioonid liidetakse kuuest reaktsioonist koosnevateks tsükliteks ning iga tsükli lõppemise tulemusena pikeneb tulevase molekuli süsinikuahel kahe süsinikuaatomi võrra.
Mõelge rasvhapete sünteesi esimeses tsüklis toimuvatele reaktsioonidele.
Esimeses kahes reaktsioonis kantakse atsetüül- ja malonüülfragmendid üle atsüüli kandvale valgule (ACP).
APB on polüpeptiidahel, mis koosneb 77 aminohappejäägist ja külgharust, mis sisuliselt kordab koensüüm A struktuuri:
Atsetüül- ja malonüülfragmentide ülekandereaktsioone atsetüül-CoA-ga (1) ja malonüül-CoA-ga (2) katalüüsivad atsüültransferaasid.
Kolmas reaktsioon seisneb atsetoatsetüül-APB moodustamises atsetüül-APB-st ja maonüül-APB-st koos samaaegse dekarboksüülimisega.
Seejärel redutseeritakse saadud atsetoatsetüül-APB kolmes etapis (reaktsioonid 4-6).
4. reaktsiooni käigus redutseeritakse üks kahest karbonüülrühmast hüdroksüüliks ja moodustub dehüdroksübuterüül-APB. See reaktsioon on NADP-st sõltuv, st. NADP redutseeritud vorm toimib redutseeriva ainena:
5. reaktsioon - dehüdratsioonireaktsioon, selle reaktsiooni ensüüm on hüdroksüatsüül-ACP dehüdraas:
Järgmine redutseerimisreaktsioon (6) - hüdrogeenimisreaktsioon - nõuab samuti NADP∙H + H + osalemist. Seda katalüüsib enoüül-APB reduktaas, reaktsiooniproduktiks on butürüül-APB:
Kõiki rasvhappeahela pikenemise (pikenemise) tsükli reaktsioone katalüüsib multiensüümide kompleks. See koosneb kahest polüpeptiidahelast. Üks neist (subühik A) sisaldab ACP-d, oksoatsüül-ACP süntaasi ja oksoatsüülreduktaasi. Subühik B sisaldab veel 4 ensüümi. Multiensüümide kompleksi koordineeritud töö on tingitud suure hoova olemasolust APB molekulis - paindlik ja üsna pikk aatomite ahel, mis ühendab "ankur" HS-rühma polüpeptiidahelaga.
Palmitiinhappe süntees hõlmab 7 tsüklit. Butürüül-APB (C 4 -atsüül) siseneb atsetüül-APB asemel teise tsüklisse ja selle tulemusena tekib kaprüül-APB (C 6 -atsüül) jne. (skeem):
1. tsükkel: malonüül-APB + atsetüül_APB
2. tsükkel: malonüül-APB + butürüül-APB
3. tsükkel: malonüül-APB + kaprüül-APB
4. tsükkel: malonüül-APB + C8-atsüül-APB
5. tsükkel: malonüül-APB + C10-atsüül-APB
6. tsükkel: malonüül-APB + C12-atsüül-APB
7. tsükkel: malonüül-APB + C14-atsüül-APB
palmitüül-APB
Palmitiinhappe biosünteesi üldvõrrand atsetüül-CoA-st seitsme tsükli reaktsioonide tulemusena on kirjutatud järgmiselt:
8 atsetüül-Coa + 7 ATP + 14 (NADP ∙ H + H +) → palmitaat + 14 NADP +
8 HS-CoA + 7 ADP + 7 H 3 RO 4
Palmitiinhappest sünteesitakse täiendavalt ühe või enama atsetüül-CoA molekuli kinnitamisel pikema ahelaga molekulid ja dehüdrogeenimisel sünteesitakse küllastumata happeid. Palmitiinhappe molekulide “viimistlemine” toimub endoplasmaatilise retikulumi ensüümide abil, kuid see võib toimuda ka mitokondrites. Küllastunud rasvhappe dehüdrogeenimine toimub paralleelselt NADP oksüdatsiooniga molekulaarse hapniku toimel:
C 15H 31 COO-S-CoA + NADP ∙ H + H + + O 2 → CH 3 - (CH 2) 5 -CH \u003d CH- (CH 2) 7 -COO-S-CoA + NADP + + 2 H2O
Küllastunud rasvhapete dehüdrogeenimine toimub maksa ja rasvkoe rakkudes. Inimkehas puuduvad ensüümid, mis võimaldaksid dehüdrogeenida -CH 2 -CH 2 - fragmente, mis asuvad C9-st kaugemal, seega dieenlinoolhape
C 18 H 32 COOH ja trieenlinoleenhape C 18 H 30 COOH organismis ei sünteesita.
Rasvhapete biosüntees toimub kõige aktiivsemalt maksarakkude, soolte, rasvkoe tsütosoolis. puhata või peale sööki.
Tavaliselt võib eristada 4 biosünteesi etappi:
1. Atsetüül-SCoA moodustumine glükoosist, teistest monosahhariididest või ketogeensetest aminohapetest.
2. Atsetüül-SCoA ülekandmine mitokondritest tsütosooli:
- võib kombineerida karnitiin, nii nagu kõrgemad rasvhapped kanduvad mitokondritesse, kuid siin toimub transport teises suunas,
- tavaliselt sisaldub sidrunhape tekkis esimeses CTC reaktsioonis.
Mitokondritest pärinev tsitraat lõhustatakse tsütosoolis ATP tsitraatlüaas oksaloatsetaadiks ja atsetüül-SCoA-ks.
Atsetüül-SCoA moodustumine sidrunhappest
Oksaloatsetaat redutseeritakse edasi malaadiks ja viimane kas siseneb mitokondritesse (malaat-aspartaadi süstik) või dekarboksüleeritakse õunensüümi ("õunhappe" ensüüm) toimel püruvaadiks.
3. Malonüül-SCoA moodustumine atsetüül-SCoA-st.
Atsetüül-SCoA karboksüülimist katalüüsib atsetüül-SCoA karboksülaas, kolme ensüümi multiensüümne kompleks.

Malonüül-SCoA moodustumine atsetüül-SCoA-st
4. Palmitiinhappe süntees.
Rakendatud multiensümaatiline kompleks" rasvhapete süntaas" (sünonüüm palmitaadi süntaas), mis sisaldab 6 ensüümi ja atsüüli kandvat valku (ACP).
Atsüüli kandev valk sisaldab pantoteenhappe derivaati - 6-fosfopanteteiin(FP), millel on HS-rühm, nagu HS-CoA. Üks kompleksi ensüüme, 3-ketoatsüülsüntaas, sisaldab ka tsüsteiini koostises HS-rühma. Nende rühmade koostoime määrab rasvhapete, nimelt palmitiinhappe biosünteesi alguse ja jätkumise. Sünteesireaktsioonid nõuavad NADPH-d.

Rasvhapete süntaasi aktiivsed rühmad
Esimeses kahes reaktsioonis seotakse malonüül-SCoA järjestikku atsüüli kandva valgu fosfopanteteiiniga ja atsetüül-SCoA 3-ketoatsüülsüntaasi tsüsteiiniga.
3-ketoatsüülsüntaas katalüüsib kolmandat reaktsiooni – atsetüülrühma üleminekut C2 malonüüliks koos karboksüülrühma elimineerimisega.
Lisaks ketorühm redutseerimisreaktsioonides ( 3-ketoatsüülreduktaas), dehüdratsioon (dehüdrataas) ja taas taastumine (enoüülreduktaas) muutub metüleeniks, moodustades küllastunud atsüüli, seotud fosfopanteteiiniga.
Atsüültransferaas kannab saadud atsüüli üle tsüsteiiniks 3-ketoatsüüli süntaasid, malonüül-SCoA kinnitatakse fosfopanteteiiniga ja tsüklit korratakse 7 korda, kuni moodustub palmitiinhappe jääk. Seejärel lõhustatakse palmitiinhape kompleksi kuuenda ensüümi tioesteraasi toimel.

Rasvhapete sünteesi reaktsioonid
Rasvhapete ahela pikenemine
Sünteesitud palmitiinhape siseneb vajadusel endoplasmaatilisesse retikulumi. Siin koos malonüül-S-CoA ja NADPH ahel pikeneb kuni C 18 või C 20 .
Ka küllastumata rasvhapped (oleiin-, linool-, linoleenhape) võivad eikosaanhappe derivaatide (C 20) moodustumisega pikeneda. Kuid kaksiksideme toovad sisse loomarakud mitte rohkem kui 9 süsinikuaatomit, seetõttu sünteesitakse ω3- ja ω6-polüküllastumata rasvhappeid ainult vastavatest lähteainetest.
Näiteks arahhidoonhape võib rakus tekkida ainult linoleen- või linoolhapete juuresolekul. Sel juhul dehüdrogeenitakse linoolhape (18:2) γ-linoleenhappeks (18:3) ja pikenetakse eikosotrieenhappeks (20:3), viimane dehüdrogeenitakse edasi arahhidoonhappeks (20:4). Nii tekivad ω6-seeria rasvhapped
ω3-seeria rasvhapete, näiteks timnodoonhappe (20:5) moodustamiseks on vajalik α-linoleenhappe (18:3) olemasolu, mis on dehüdreeritud (18:4), pikenenud (20:4). ) ja dehüdreeriti uuesti (20:5).
Atsetüül-CoA moodustumine ja selle transport tsütosooli
Rasvhapete süntees toimub imendumisperioodil. Püruvaadi aktiivne glükolüüs ja sellele järgnev oksüdatiivne dekarboksüülimine aitavad kaasa atsetüül-CoA kontsentratsiooni suurenemisele mitokondriaalses maatriksis. Kuna rasvhapete süntees toimub rakkude tsütosoolis, tuleb atsetüül-CoA transportida läbi sisemise mitokondriaalse membraani tsütosooli. Sisemine mitokondriaalne membraan on aga atsetüül-CoA-le mitteläbilaskev, seetõttu kondenseerub atsetüül-CoA mitokondriaalses maatriksis oksaloatsetaadiga, moodustades tsitraadi tsitraadi süntaasi osalusel:
Atsetüül-CoA + oksaloatsetaat -> tsitraat + HS-CoA.
Seejärel transpordib translokaas tsitraadi tsütoplasmasse (joonis 8-35).
Tsitraadi ülekanne tsütoplasmasse toimub ainult tsitraadi koguse suurenemisega mitokondrites, kui isotsitraadi dehüdrogenaasi ja α-ketoglutaraadi dehüdrogenaasi inhibeerivad kõrged NADH ja ATP kontsentratsioonid. Selline olukord tekib imendumisperioodil, mil maksarakk saab piisavas koguses energiaallikaid. Tsütoplasmas lõhustatakse tsitraat ensüümi tsitraatlüaasi toimel:
Tsitraat + HSKoA + ATP → atsetüül-CoA + ADP + Pi + oksaloatsetaat.
Tsütoplasmas olev atsetüül-CoA toimib rasvhapete sünteesi algse substraadina ja tsütosoolis olev oksaloatsetaat läbib järgmised transformatsioonid (vt allolevat diagrammi).
Püruvaat transporditakse tagasi mitokondriaalsesse maatriksisse. Maleiinensüümi toime tulemusena redutseeritud NADPH-d kasutatakse vesiniku doonorina järgmistes rasvhapete sünteesi reaktsioonides. Teine NADPH allikas on glükoosi katabolismi pentoosfosfaadi raja oksüdatiivsed etapid.
Malonüül-CoA moodustumine atsetüül-CoA-st - rasvhapete biosünteesi reguleeriv reaktsioon.
Esimene reaktsioon rasvhapete sünteesil on atsetüül-CoA muundamine malonüül-CoA-ks. Seda reaktsiooni katalüüsiv ensüüm (atsetüül-CoA karboksülaas) kuulub ligaaside klassi. See sisaldab kovalentselt seotud biotiini (joonis 8-36). Reaktsiooni esimeses etapis seostub CO 2 ATP energia toimel kovalentselt biotiiniga, teises etapis kantakse COO atsetüül-CoA-ks koos malonüül-CoA moodustumisega. Atsetüül-CoA karboksülaasi ensüümi aktiivsus määrab kõigi järgnevate rasvhapete sünteesireaktsioonide kiiruse.
Rasvhapete süntaasi katalüüsitud reaktsioonid- ensüümikompleks, mis katalüüsib palmitiinhappe sünteesi reaktsioone, on kirjeldatud allpool.
Pärast malonüül-CoA moodustumist jätkub rasvhapete süntees multiensüümikompleksil – rasvhapete süntaasil (palmitoüülsüntetaas). See ensüüm koosneb 2 identsest protomeerist, millest igaühel on domeeni struktuur ja vastavalt 7 erineva katalüütilise aktiivsusega tsentrit (joonis 8-37). See kompleks pikendab järjestikku 2 süsinikuaatomi võrra rasvhapperadikaali, mille doonoriks on malonüül-CoA. Selle kompleksi lõpp-produkt on palmitiinhape, seega on selle ensüümi endine nimetus palmitoüülsüntetaas.
Esimene reaktsioon on atsetüül-CoA atsetüülrühma ülekandmine tsüsteiini tioolrühmale atsetüültransatsülaasi tsentri poolt (joonis 8-38). Malonüüli jääk viiakse seejärel malonüül-CoA-st atsüüli kandva valgu sulfhüdrüülrühma malonüültransatsülaasi tsentri abil. Pärast seda on kompleks valmis esimeseks sünteesitsükliks.
Atsetüülrühm kondenseerub ülejäänud malonüüliga eraldatud CO 2 kohas. Reaktsiooni katalüüsib ketoatsüülsüntaasi tsenter. Saadud atsetoatsetüülradikaal
Skeem

Riis. 8-35. Atsetüüli jääkide ülekandmine mitokondritest tsütosooli. Aktiivsed ensüümid: 1 - tsitraadi süntaas; 2 - translokaas; 3 - tsitraatlüaas; 4 - malaatdehüdrogenaas; 5 - malik-ensüüm.

Riis. 8-36. Biotiini roll atsetüül-CoA karboksüülimise reaktsioonis.

Riis. 8-37. Multiensüümide kompleksi struktuur on rasvhapete süntees. Kompleks on kahe identse polüpeptiidahela dimeer, millest igaühel on 7 aktiivset saiti ja atsüüli kandev valk (ACP). Protomeeride SH rühmad kuuluvad erinevatesse radikaalidesse. Üks SH rühm kuulub tsüsteiini, teine fosfopanteethappe jäägi hulka. Ühe monomeeri tsüsteiini-SH-rühm asub teise protomeeri 4-fosfopanteteinaat-SH-rühma kõrval. Seega on ensüümi protomeerid paigutatud peast-saba. Kuigi iga monomeer sisaldab kõiki katalüütilisi saite, on kahest protomeerist koosnev kompleks funktsionaalselt aktiivne. Seetõttu sünteesitakse tegelikult korraga 2 rasvhapet. Lihtsuse huvides on skeemidel tavaliselt kujutatud reaktsioonide jada ühe happemolekuli sünteesil.
redutseeritakse järjestikku ketoatsüülreduktaasi poolt, seejärel dehüdreeritakse ja taas redutseeritakse kompleksi aktiivsete tsentrite enoüülreduktaasi poolt. Esimese reaktsioonitsükli tulemusena moodustub butürüülradikaal, mis on seotud rasvhapete süntaasi subühikuga.
Enne teist tsüklit viiakse butürüülradikaal positsioonilt 2 positsioonile 1 (kus atsetüül asus esimese reaktsioonitsükli alguses). Seejärel läbib butürüüli jääk samasugused muutused ja pikeneb 2 süsinikuaatomi võrra, mis pärineb malonüül-CoA-st.
Sarnaseid reaktsioonide tsükleid korratakse, kuni moodustub palmitiinhapperadikaal, mis tioesteraasi tsentri toimel eraldub hüdrolüütiliselt ensüümikompleksist, muutudes vabaks palmitiinhappeks (palmitaat, joon. 8-38, 8-39).
Palmitiinhappe atsetüül-CoA ja malonüül-CoA sünteesi üldvõrrand on järgmine:
CH 3 -CO-SKoA + 7 HOOC-CH 2 -CO-SKoA + 14 (NADPH + H+) → C 15 H 31 COOH + 7 CO 2 + 6 H 2 O + 8 HSKoA + 14 NADP +.
Peamised vesiniku allikad rasvhapete sünteesiks
Igas palmitiinhappe biosünteesi tsüklis toimub 2 redutseerimisreaktsiooni,

Riis. 8-38. Palmitiinhappe süntees. Rasvhapete süntaas: esimeses protomeeris kuulub SH-rühm tsüsteiinile, teises fosfopanteteiinile. Pärast esimese tsükli lõppu kantakse butürüülradikaal üle esimese protomeeri SH-rühma. Seejärel korratakse sama reaktsioonide jada nagu esimeses tsüklis. Palmitoüül-E on palmitiinhappe jääk, mis on seotud rasvhapete süntaasiga. Sünteesitud rasvhappes pärinevad atsetüül-CoA-st ainult 2 distaalset süsinikku, mis on tähistatud *-ga, ülejäänud malonüül-CoA-st.

Riis. 8-39. Üldskeem palmitiinhappe sünteesi reaktsioonid.
milles koensüüm NADPH toimib vesiniku doonorina. NADP + taastumine toimub reaktsioonides:
dehüdrogeenimine glükoosi katabolismi pentoosfosfaadi raja oksüdatiivsetes etappides;
malaadi dehüdrogeenimine õunensüümiga;
isotsitraadi dehüdrogeenimine tsütosoolse NADP-sõltuva dehüdrogenaasi poolt.
2. Rasvhapete sünteesi reguleerimine
Rasvhapete sünteesi reguleeriv ensüüm on atsetüül-CoA karboksülaas. Seda ensüümi reguleeritakse mitmel viisil.
Ensüümide alaühikute komplekside assotsiatsioon/dissotsiatsioon. Inaktiivsel kujul on atsetüül-CoA karboksülaas eraldi kompleks, millest igaüks koosneb 4 alaühikust. Ensüümi aktivaator - tsitraat; see stimuleerib komplekside seostumist, mille tulemusena ensüümi aktiivsus suureneb. Inhibiitor - palmitoüül-CoA; see põhjustab kompleksi dissotsiatsiooni ja ensüümi aktiivsuse vähenemist (joon. 8-40).
Atsetüül-CoA karboksülaasi fosforüülimine/defosforüülimine. Postabsorptiivses seisundis või füüsilise töö ajal aktiveerib glükagoon või adrenaliin adenülaattsüklaasi süsteemi kaudu proteiinkinaasi A ja stimuleerib atsetüül-CoA karboksülaasi subühikute fosforüülimist. Fosforüülitud ensüüm on inaktiivne ja rasvhapete süntees peatub. Imendumisperioodil aktiveerib insuliin fosfataasi ja atsetüül-CoA karboksülaas defosforüleerub (joonis 8-41). Seejärel toimub tsitraadi toimel ensüümi protomeeride polümerisatsioon ja see muutub aktiivseks. Tsitraadil on lisaks ensüümi aktiveerimisele ka teine funktsioon rasvhapete sünteesis. Imendumisperioodil koguneb maksarakkude mitokondritesse tsitraat, milles atsetüüli jääk transporditakse tsütosooli.
Ensüümide sünteesi indutseerimine. Süsivesikuterikka ja rasvavaese toidu pikaajaline tarbimine põhjustab insuliini sekretsiooni suurenemist, mis stimuleerib ensüümide sünteesi indutseerimist: atsetüül-CoA karboksülaas, rasvhapete süntaas, tsitraatlüaas,

Riis. 8-40. Atsetüül-CoA karboksülaasi komplekside assotsiatsioon/dissotsiatsioon.

Riis. 8-41. Atsetüül-CoA karboksülaasi reguleerimine.
Riis. 8-42. Palmitiinhappe pikenemine ER-s. Palmitiinhappe radikaal on pikenenud 2 süsinikuaatomi võrra, mille doonoriks on malonüül-CoA.
isotsitraatdehüdrogenaas. Seetõttu kiirendab süsivesikute liigne tarbimine glükoosi katabolismi produktide muundumist rasvadeks. Nälgimine või rasvarikas toit põhjustab ensüümide ja vastavalt ka rasvade sünteesi vähenemist.
3. Rasvhapete süntees palmitiinhappest
Rasvhapete pikenemine. ER-s pikendatakse palmitiinhapet malonüül-CoA osalusel. Reaktsioonide jada on sarnane palmitiinhappe sünteesi ajal toimuvaga, kuid sel juhul seostatakse rasvhappeid mitte rasvhapete süntaasi, vaid CoA-ga. Elongatsioonis osalevad ensüümid võivad substraatidena kasutada mitte ainult palmitiinhapet, vaid ka teisi rasvhappeid (joonis 8-42), seetõttu saab organismis sünteesida mitte ainult steariinhapet, vaid ka suure süsinikuaatomite arvuga rasvhappeid.
Peamine pikenemise produkt maksas on steariinhape (С 18:0), kuid ajukoes tekib see suur hulk pikema ahelaga rasvhapped - C 20 kuni C 24, mis on vajalikud sfingolipiidide ja glükolipiidide moodustamiseks.
Närvikoes toimub ka teiste rasvhapete, α-hüdroksühapete süntees. oksüdaas co segafunktsioonid hüdroksülaadid C 22 ja C 24 happed koos lignotseriini ja tserebroonhappe moodustumisega, mida leidub ainult aju lipiidides.
Kaksiksidemete moodustumine rasvhapperadikaalides. Kaksiksidemete liitumist rasvhapperadikaalidega nimetatakse desaturatsiooniks. Peamised rasvhapped, mis inimorganismis tekivad desaturatsiooni tulemusena (joonis 8-43), on palmitooleiinhape (C16:1Δ9) ja oleiinhape (C18:1Δ9).
Kaksiksideme moodustumine rasvhapperadikaalides toimub ER-s reaktsioonides, mis hõlmavad molekulaarset hapnikku, NADH-d ja tsütokroom b 5 . Inimorganismis esinevad rasvhapete desaturaasi ensüümid ei saa moodustada kaksiksidet rasvhapperadikaalides, mis asuvad üheksandast süsinikuaatomist kaugemal, s.o. üheksanda ja vahel

Riis. 8-43. Küllastumata rasvhapete moodustumine.
metüül süsiniku aatomid. Seetõttu ei sünteesita ω-3 ja ω-6 perekonda kuuluvaid rasvhappeid organismis, need on asendamatud ja neid tuleb varustada toiduga, kuna nad täidavad olulisi reguleerivaid funktsioone.
Kaksiksideme moodustamiseks rasvhapperadikaalis on vaja molekulaarset hapnikku, NADH-d, tsütokroom b 5 ja FAD-sõltuvat tsütokroom b 5 reduktaasi. Küllastunud happest eraldatud vesinikuaatomid vabanevad veena. Üks molekulaarne hapnikuaatom sisaldub veemolekulis ja teine redutseeritakse samuti veeks NADH elektronide osalusel, mis kanduvad üle FADH 2 ja tsütokroom b 5 kaudu.
Eikosanoidid on bioloogiliselt aktiivsed ained, mida enamik rakke sünteesivad 20 süsinikuaatomit sisaldavatest polüeenrasvhapetest (sõna "eikosa" tähendab kreeka keeles 20).
Atsetüül-CoA on VFA-de sünteesi substraat, kuid rasvhapete (FA) sünteesi käigus ei kasutata igas elongatsioonitsüklis mitte atsetüül-CoA-d ennast, vaid selle derivaati malonüül-CoA-d.
Seda reaktsiooni katalüüsib ensüüm atsetüül-CoA karboksülaas, mis on FA sünteesi multiensüümsüsteemi võtmeensüüm. Ensüümi aktiivsust reguleerib negatiivse tagasiside tüüp. Inhibiitoriks on sünteesiprodukt: pika ahelaga atsüül-CoA (n=16) - palmitoüül-CoA. Aktivaatoriks on tsitraat. Selle ensüümi mittevalguline osa sisaldab H-vitamiini (biotiini).
Seejärel pikeneb atsüül-CoA molekul rasvhapete sünteesi käigus iga etapi kohta järk-järgult 2 süsinikuaatomi võrra malonüül-CoA tõttu, mis kaotab selles pikenemisprotsessis CO 2.
Pärast malonüül-CoA moodustumist katalüüsib rasvhapete sünteesi peamisi reaktsioone üks ensüüm - rasvhapete süntetaas (kinnitub endoplasmaatilise retikulumi membraanidele). Rasvhapete süntetaas sisaldab 7 aktiivset saiti ja atsüüli kandvat valku (ACP). Malonüül-CoA sidumissait sisaldab mittevalgukomponenti, vitamiini B3 (pantoteenhape). Ühe reaktsioonitsükli järjestus HFA sünteesiks on näidatud joonisel 45.





Joonis 45. Kõrgemate rasvhapete sünteesi reaktsioonid
Pärast tsükli lõppu siseneb atsüül-APB järgmisse sünteesitsüklisse. Atsüüli kandva valgu vaba SH-rühma külge on kinnitatud uus malonüül-CoA molekul. Seejärel eraldatakse atsüüljääk, see kantakse üle malonüüljäägile (samaaegse dekarboksüülimisega) ja reaktsioonitsüklit korratakse.
Seega kasvab tulevase rasvhappe süsivesinike ahel järk-järgult (iga tsükli kohta kahe süsinikuaatomi võrra). See juhtub seni, kuni see pikeneb 16 süsinikuaatomini (palmitiinhappe sünteesi korral) või rohkem (teiste rasvhapete süntees). Pärast seda toimub tiolüüs ja rasvhappe aktiivne vorm atsüül-CoA moodustub valmis kujul.
Kõrgemate rasvhapete sünteesi normaalseks kulgemiseks on vajalikud järgmised tingimused:
1) Süsivesikute tarbimine, mille oksüdatsiooni käigus tekivad vajalikud substraadid ja NADPH 2.
2) Raku kõrge energialaeng - kõrge ATP sisaldus, mis tagab tsitraadi vabanemise mitokondritest tsütoplasmasse.
Võrdlevad omadused b-oksüdatsioon ja kõrgemate rasvhapete süntees:
1 . b-oksüdatsioon toimub mitokondrites ja rasvhapete süntees toimub tsütoplasmas endoplasmaatilise retikulumi membraanidel. Mitokondrites moodustunud atsetüül-CoA ei saa aga ise membraane läbida. Seetõttu on olemas mehhanismid atsetüül-CoA transportimiseks mitokondritest tsütoplasmasse Krebsi tsükli ensüümide osalusel (joonis 46).

Joonis 46. Atsetüül-CoA transportimise mehhanism mitokondritest tsütoplasmasse.
TCA peamised ensüümid on tsitraadi süntaas ja isotsitraatdehüdrogenaas. Nende ensüümide peamised allosteerilised regulaatorid on ATP ja ADP. Kui rakus on palju ATP-d, toimib ATP nende võtmeensüümide inhibiitorina. Siiski inhibeerib ATP isotsitraatdehüdrogenaasi rohkem kui tsitraadi süntetaasi. See viib tsitraadi ja isotsitraadi kuhjumiseni mitokondriaalses maatriksis. Kogunemisel lahkub tsitraat mitokondritest ja siseneb tsütoplasmasse. Tsütoplasma sisaldab ensüümi tsitraatlüaasi. See ensüüm lagundab tsitraadi PAA-ks ja atsetüül-CoA-ks.
Seega on atsetüül-CoA mitokondritest tsütoplasmasse vabanemise tingimuseks raku hea varustamine ATP-ga. Kui rakus on vähe ATP-d, lõhustatakse atsetüül-CoA CO 2 ja H 2 O-ks.
2 . B-oksüdatsiooni käigus seostuvad vaheühendid HS-CoA-ga ning rasvhapete sünteesi käigus vaheühendid spetsiifilise atsüüli kandva valguga (ACP). See on kompleksne valk. Selle mittevalguline osa on struktuurilt sarnane CoA-ga ja koosneb tioetüülamiinist, pantoteenhappest (vitamiin B 3) ja fosfaadist.
3 . B-oksüdeerimisel kasutatakse oksüdeerijana NAD ja FAD. Rasvhapete sünteesil on vaja redutseerivat ainet - kasutatakse NADP * H 2.
Rasvhapete sünteesiks on rakus kaks peamist NADP * H 2 allikat:
a) süsivesikute lagunemise pentoosfosfaadi rada;
Rasvhapete sünteesi ehitusplokk raku tsütosoolis on atsetüül-CoA, mis tekib kahel viisil: kas püruvaadi oksüdatiivse dekarboksüülimise tulemusena. (vt joonis 11, III etapp), või rasvhapete b-oksüdatsiooni tulemusena (vt joonis 8).
Joonis 11 – süsivesikute lipiidideks muundamise skeem
Tuletame meelde, et glükolüüsi käigus moodustunud püruvaadi muundumine atsetüül-CoA-ks ja selle moodustumine rasvhapete b-oksüdatsiooni käigus toimub mitokondrites. Rasvhapete süntees toimub tsütoplasmas. Mitokondrite sisemembraan on atsetüül-CoA-le mitteläbilaskev. Selle sisenemine tsütoplasmasse toimub hõlbustatud difusiooni tüübi abil tsitraadi või atsetüülkarnitiini kujul, mis tsütoplasmas muundatakse atsetüül-CoA-ks, oksaloatsetaadiks või karnitiiniks. Kuid peamine viis atsetüül-coA ülekandmiseks mitokondritest tsütosooli on tsitraat (vt joonis 12).
Esialgu interakteerub intramitokondriaalne atsetüül-CoA oksaloatsetaadiga, mille tulemusena moodustub tsitraat. Reaktsiooni katalüüsib ensüüm tsitraadi süntaas. Saadud tsitraat transporditakse spetsiaalse trikarboksülaadi transpordisüsteemi abil läbi mitokondriaalse membraani tsütosooli.
Tsütosoolis reageerib tsitraat HS-CoA ja ATP-ga, lagunedes uuesti atsetüül-CoA-ks ja oksaloatsetaadiks. Seda reaktsiooni katalüüsib ATP-tsitraatlüaas. Juba tsütosoolis naaseb oksaloatsetaat tsütosoolse dikarboksülaati transportiva süsteemi osalusel mitokondriaalsesse maatriksisse, kus see oksüdeeritakse oksaloatsetaadiks, viies sellega lõpule nn süstikutsükli:
Joonis 12 – atsetüül-CoA mitokondritest tsütosooli ülekande skeem
Küllastunud rasvhapete biosüntees toimub nende b-oksüdatsioonile vastupidises suunas, rasvhapete süsivesinike ahelate kasv toimub kahe süsiniku fragmendi (C 2) - atsetüül-CoA järjestikuse lisamise tõttu nende otstesse. (vt joonis 11, etapp IV).
Rasvhapete biosünteesi esimene reaktsioon on atsetüül-CoA karboksüülimine, milleks on vaja CO 2, ATP, Mn ioone. Seda reaktsiooni katalüüsib ensüüm atsetüül-CoA-karboksülaas. Ensüüm sisaldab proteesrühmana biotiini (H-vitamiini). Reaktsioon toimub kahes etapis: 1 - biotiini karboksüülimine ATP ja II osalusel - karboksüülrühma ülekandmine atsetüül-CoA-le, mille tulemusena moodustub malonüül-CoA:
Malonüül-CoA on esimene spetsiifiline rasvhapete biosünteesi produkt. Sobiva ensüümsüsteemi juuresolekul muundub malonüül-CoA kiiresti rasvhapeteks.
Tuleb märkida, et rasvhapete biosünteesi kiiruse määrab suhkrute sisaldus rakus. Glükoosi kontsentratsiooni suurenemine inimeste ja loomade rasvkoes ja glükolüüsi kiiruse suurenemine stimuleerivad rasvhapete sünteesi. See näitab, et rasvade ja süsivesikute ainevahetus on omavahel tihedalt seotud. Olulist rolli mängib siin atsetüül-CoA karboksüülimise reaktsioon selle muundumisega malonüül-CoA-ks, mida katalüüsib atsetüül-CoA karboksülaas. Viimaste aktiivsus sõltub kahest tegurist: suure molekulmassiga rasvhapete ja tsitraadi olemasolust tsütoplasmas.
Rasvhapete akumuleerumine pärsib nende biosünteesi; pärsivad karboksülaasi aktiivsust.
Eriline roll on tsitraadil, mis on atsetüül-CoA karboksülaasi aktivaator. Tsitraat mängib samal ajal lüli rolli süsivesikute ja rasvade ainevahetus. Tsütoplasmas on tsitraadil rasvhapete sünteesi stimuleerimisel kahekordne toime: esiteks atsetüül-CoA karboksülaasi aktivaatorina ja teiseks atsetüülrühmade allikana.
Rasvhapete sünteesi väga oluline tunnus on see, et kõik sünteesi vaheühendid on kovalentselt seotud atsüülkandjavalguga (HS-ACP).
HS-ACP on madala molekulmassiga valk, mis on termostabiilne, sisaldab aktiivset HS-rühma ja mille proteesrühmas on pantoteenhape (vitamiin B3). HS-ACP funktsioon on sarnane ensüümi A (HS-CoA) funktsiooniga rasvhapete b-oksüdeerimisel.
Rasvhappeahela ehitamisel moodustavad vaheühendid estersidemeid loomsete kõrvalsaaduste ahelaga (vt joonis 14):
Rasvhappeahela pikenemise tsükkel sisaldab nelja reaktsiooni: 1) atsetüül-APB (C2) kondenseerimine malonüül-APB-ga (C3); 2) taastumine; 3) dehüdratsioon ja 4) rasvhapete teine taastumine. Joonisel fig. 13 on näidatud rasvhapete sünteesi skeem. Üks rasvhappeahela pikendamise tsükkel hõlmab nelja järjestikust reaktsiooni.
Joonis 13 – Rasvhapete sünteesi skeem
Esimeses reaktsioonis (1) - kondensatsioonireaktsioonis - interakteeruvad atsetüül- ja malonüülrühmad üksteisega, moodustades atsetoatsetüül-ABP koos samaaegse CO 2 (C 1) vabanemisega. Seda reaktsiooni katalüüsib kondenseeruv ensüüm b-ketoatsüül-ABP süntetaas. Malonüül-APB-st lõhustatud CO2 on sama CO2, mis osales atsetüül-APB karboksüülimisreaktsioonis. Seega tekib kondensatsioonireaktsiooni tulemusena kahe (C 2) ja kolme süsiniku (C 3) komponendist neljasüsinikuline ühend (C 4).
Teises reaktsioonis (2), redutseerimisreaktsioon, mida katalüüsib b-ketoatsüül-ACP reduktaas, atsetoatsetüül-ACB muundatakse b-hüdroksübutürüül-ACB-ks. Redutseerija on NADPH + H +.
Dehüdratsioonitsükli kolmandas reaktsioonis (3) eraldatakse veemolekul b-hüdroksübutürüül-APB-st, moodustades krotonüül-APB. Reaktsiooni katalüüsib b-hüdroksüatsüül-ACP dehüdraas.
Tsükli neljas (viimane) reaktsioon (4) on krotoniil-APB redutseerimine butürüül-APB-ks. Reaktsioon kulgeb enoüül-ACP reduktaasi toimel. Redutseerija rolli täidab siin teine molekul NADPH + H + .
Seejärel korratakse reaktsioonide tsüklit. Oletame, et palmitiinhapet (C 16) sünteesitakse. Sel juhul viiakse butürüül-ACB moodustumine lõpule alles esimese 7-st tsüklist, millest igaühe algus on molonüül-ACB molekuli (C 3) lisamine - reaktsioon (5) karboksüüli lõppu. kasvav rasvhappeahel. Sel juhul eraldatakse karboksüülrühm CO 2 (C1) kujul. Seda protsessi saab kujutada järgmiselt:
C 3 + C 2 ® C 4 + C 1 - 1 tsükkel
C 4 + C 3 ® C 6 + C 1 - 2 tsükkel
C 6 + C 3 ® C 8 + C 1 -3 tsükkel
C 8 + C 3 ® C 10 + C 1-4 tsükkel
C 10 + C 3 ® C 12 + C 1-5 tsükkel
C 12 + C 3 ® C 14 + C 1-6 tsükkel
C 14 + C 3 ® C 16 + C 1-7 tsükkel
Sünteesida saab mitte ainult kõrgemaid küllastunud rasvhappeid, vaid ka küllastumata rasvhappeid. Monoküllastumata rasvhapped tekivad küllastunud rasvhapetest oksüdatsiooni (desaturatsiooni) tulemusena, mida katalüüsib atsüül-CoA oksügenaas. Erinevalt taimsetest kudedest on loomsetel kudedel väga piiratud võime muuta küllastunud rasvhappeid küllastumata rasvhapeteks. On kindlaks tehtud, et kaks levinumat monoküllastumata rasvhapet, palmitooleiin- ja oleiinhape, sünteesitakse palmitiin- ja steariinhappest. Imetajate, sealhulgas inimeste kehas ei saa steariinhappest (C 18:0) tekkida näiteks linoolhape (C 18:2) ja linoleenhape (C 18:3). Need happed on klassifitseeritud asendamatuteks rasvhapeteks. Asendamatute rasvhapete hulka kuulub ka arahhiidhape (C 20:4).
Koos rasvhapete desaturatsiooniga (kaksiksidemete moodustumine) toimub ka nende pikenemine (pikenemine). Lisaks saab neid mõlemaid protsesse kombineerida ja korrata. Rasvhappeahela pikenemine toimub kahe süsiniku fragmendi järjestikuse lisamisega vastavale atsüül-CoA-le malonüül-CoA ja NADPH+H+ osalusel.
Joonisel 14 on kujutatud palmitiinhappe transformatsiooniteed desaturatsiooni- ja pikenemisreaktsioonides.
Joonis 14 - Küllastunud rasvhapete muundamise skeem
küllastumatuks
Mis tahes rasvhappe süntees viiakse lõpule HS-ACP lõhustamisega atsüül-ACB-st deatsülaasi ensüümi mõjul. Näiteks:
Saadud atsüül-CoA on aktiivne vorm rasvhape.